the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 19 May 2025
| 19 May 2025
Impact of gravel bar flooding on arthropods in the upper Isar: implications for river management
Andrea Sundermann
Braided rivers are dynamic ecosystems with constantly shifting flow patterns that create diverse habitats, supporting species highly adapted to frequent flooding and gravel relocation. Arthropods inhabiting these areas use various strategies to survive floods, including migration, hiding, and physiological adaptations like flying or surviving submerged. This study aims to assess whether certain riverbank areas act as refuges during floods and to determine if dominant arthropod species actively or passively respond to rising water levels, with implications for the conservation of these ecosystems. This study focused on the spatial distribution and flood response of two spider species and two beetle species in a gravel bank area along the upper Isar (Germany). Species distributions were recorded under normal conditions and compared to their distribution after a flood event, using 203 sampling squares. In addition, an artificial flood experiment was conducted to observe species behaviour under rising water. Flooding significantly altered the distribution of the studied species, with many individuals relocating to higher areas like slopes during peak flood and scattering across the exposed gravel bar once waters receded. In an artificial flood experiment, individuals initially took refuge in gravel but eventually crossed the water's surface to escape rising water levels, showing varied survival behaviours like floating on stones or paddling to safety. The study highlights the critical role of accessible elevated areas and riverbanks as refuges for arthropods during flooding, with 39 % of individuals actively utilizing these sites in response to rising water levels. The findings emphasize that riverbanks and elevated areas must be preserved and managed effectively, as they provide vital refuge conditions for arthropod communities, ultimately fostering ecological resilience in natural river systems.
- Article
(3880 KB) - Full-text XML
-
Supplement
(531 KB) - BibTeX
- EndNote




Natural braided rivers and their floodplains are highly dynamic ecosystems (Leopold and Wolman, 1957; Tockner et al., 2006; Van der Nat et al., 2003). Their flow patterns, with multiple channels and braids, shift and change constantly over time. This creates a riverbed with varied topography, consisting of an alluvial habitat mosaic that includes all successional stages, from bare gravel to tree-covered patches (Ferguson and Ashworth, 1992; Ward et al., 1999; Ward and Tockner, 2001). Sediment load and flood pulses are the driving forces behind this dynamic (Junk, 1999; Wohl, 2014). The complex flow patterns and the constantly evolving environment are key to the ecosystem's ability to provide habitats for numerous species, most of which are highly adapted to specific conditions. The open, exposed gravel areas form a transitional zone between aquatic and terrestrial habitats. Arthropods inhabiting this zone have evolved various adaptations (Eyre and Lott, 1997; Manderbach, 1998; Framenau et al.,1996; Plachter, 1986) to cope with unpredictable flooding events (Gewässerkundlicher Dienst Bayern, 2025; Campolo et al., 1999). The species living in these harsh environments can employ different strategies to cope with flooding. Some species simply leave the floodplain area during the flood season and resettle during low-flow periods. Resettlement can be either directed or undirected (Rothenbücher and Schaefer, 2006; Weigmann and Wohlgemuth-von Reiche, 1999; Zulka, 1994a). Other strategies to cope with flooding include hiding between the gravel (Andersen, 1968; Kühnelt, 1943; Lehmann, 1965) or escaping to higher elevated areas and trees, which serve as refuge areas (Kühnelt, 1943; Manderbach, 1998; Zulka, 1994a). Physiological adaptations, such as the ability to fly (Andersen, 1968; Bonn, 2000; Desender, 1989; Kühnelt, 1943; Zulka, 1994a) or move on the water surface using surface tension (Kühnelt, 1943; Lambeets and Bonte, 2009; Manderbach and Plachter, 1997; Papi, 1955; Perevozkin et al., 2004; Wessels and Sundermann, 2022), like the two spider genera Arctosa and Pardosa (Kühnelt, 1943), increase the probability of survival. Carabidae and Staphylinidae have been observed to dive and hide under stones using large air pockets beneath their elytra (Kühnelt, 1943; Manderbach and Plachter, 1997). Dorsal ventrally flattened bodies enable Carabidae to hide between gravel (Andersen, 1968; Desender, 1989; Kühnelt, 1943), where air inclusions in unsaturated sediments allow them to survive a flood. Physiological flood resistance and the ability to survive submerged in inundated areas have been described by several authors (Andersen, 1968; Hebets and Chapman, 2000; Kühnelt, 1943; Rothenbücher and Schaefer, 2006; Weigmann and Wohlgemuth-von Reiche, 1999; Zulka, 1994a). Adaptations in life cycles have also been observed (Zulka, 1994b), such as in the spider Arctosa cinerea, which has an asynchronous life cycle, with spiderlings and adults always coexisting. Different habitats for different life stages reduce the loss of a population during flooding. In this regard, larvae of Nebria picicornis were, unlike adults, in areas away from the water's edge (Manderbach and Plachter, 1997). Nevertheless, the majority of the strategies outlined above may prove inadequate when flood events are severe enough to displace entire gravel banks. In such instances, the responses of the species remain largely unknown.
However, understanding how species respond to flood events helps identify key refuges and adaptive strategies, which is essential for conservation efforts as anthropogenic influences such as river regulation, habitat fragmentation, and climate change threaten these natural dynamics, potentially reducing the ability of these systems to support biodiversity. By studying the interactions between species and their environment during flood events, researchers can better inform management strategies aimed at preserving the ecological balance of these systems. Protecting braided rivers not only ensures the survival of the species adapted to these unique habitats but also helps maintain biodiversity on a broader scale, contributing to the resilience of ecosystems and their capacity to provide essential services. In this context, the present study aims to assess, in braided river systems, (1) to which degree riverbanks and elevated areas within the riverbed function as refuges during flood events and (2) to which degree the dominant arthropod species within the studied river section exhibit active or passive responses to rising water levels.
2.1 Studied species and study area
This study focused on two spider species, Arctosa cinerea and Pardosa wagleri, as well as the rove beetle Paederidus rubrothoracicus, within the upper Isar between river kilometre 233.8 and 234.1 near Vorderriß (Fig. 1). These three species were identified as the most abundant riparian arthropod species in this area (Wessels and Sundermann, 2022). Additionally, the ground beetle Nebria picicornis and several species of the genus Bembidion were included in the study, as they have been classified by other authors as typical gravel bar inhabitants (Manderbach, 2002). Since species-level identification within Bembidion often requires binocular magnification, this genus was recorded collectively without distinguishing individual species. Field studies were conducted in the summer of 2014 on a gravel bank located between the southern riverbank and a primary water channel. The riverbank consists of a steep slope approximately 10 m high. At the ends of the gravel bar, the slope transitions into an almost vertical drop-off edge. The vertical drop-off edge, along with the adjacent forest area, separates the gravel bank from other gravel habitats, making the migration of gravel bank arthropods via this route unlikely. Due to the relatively recent formation of the gravel bank – estimated to be no older than 2 years based on accounts from local residents, aerial photographs from 2012, and structural comparisons with conditions documented in 2011 – vegetation development has not yet commenced.
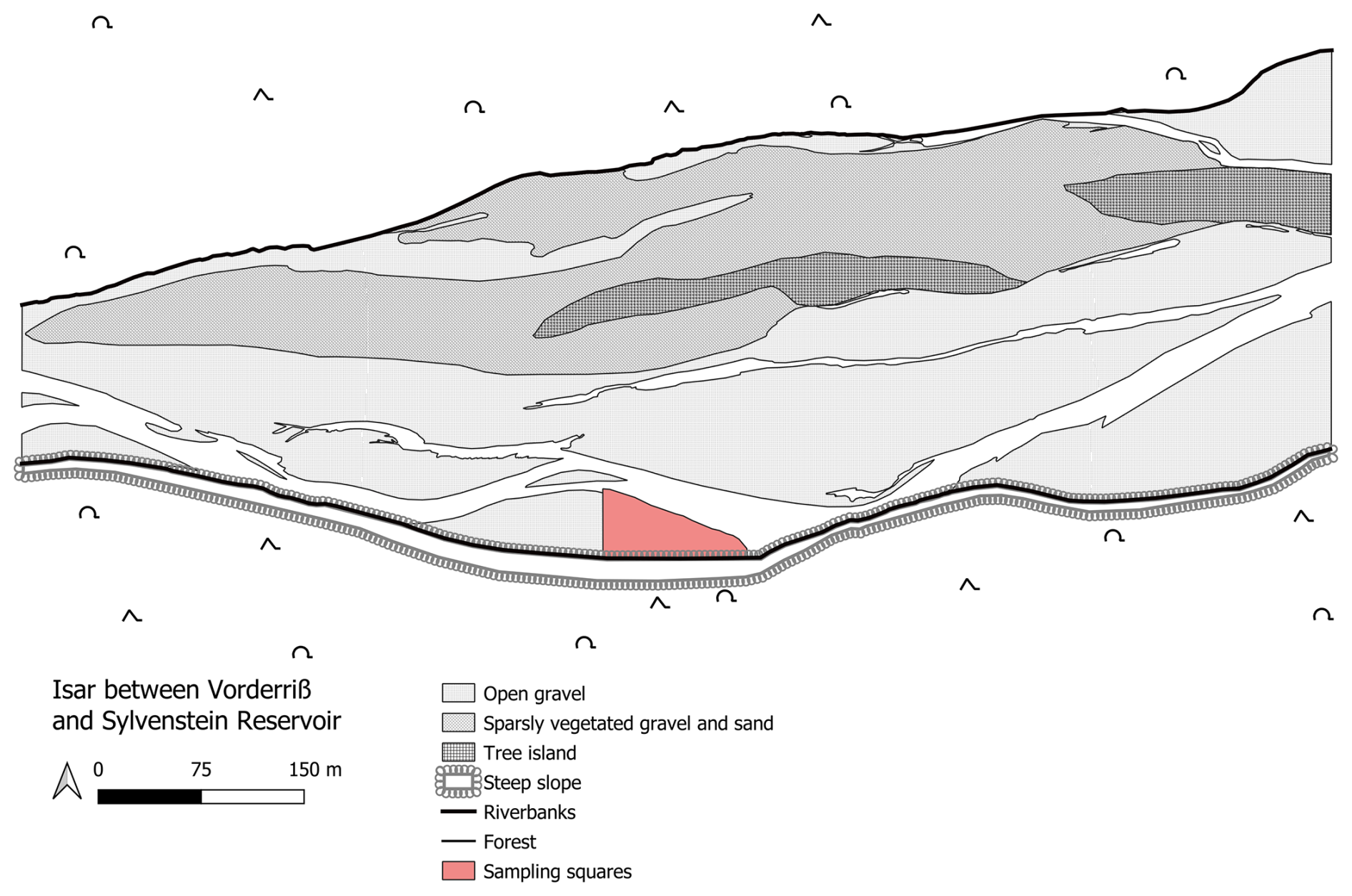
Figure 1River stretch including the sampled gravel bank (red). Flow direction from left to right.

Figure 2View from north to the gravel bank, with the study area (framed in red) and the slope in the background.
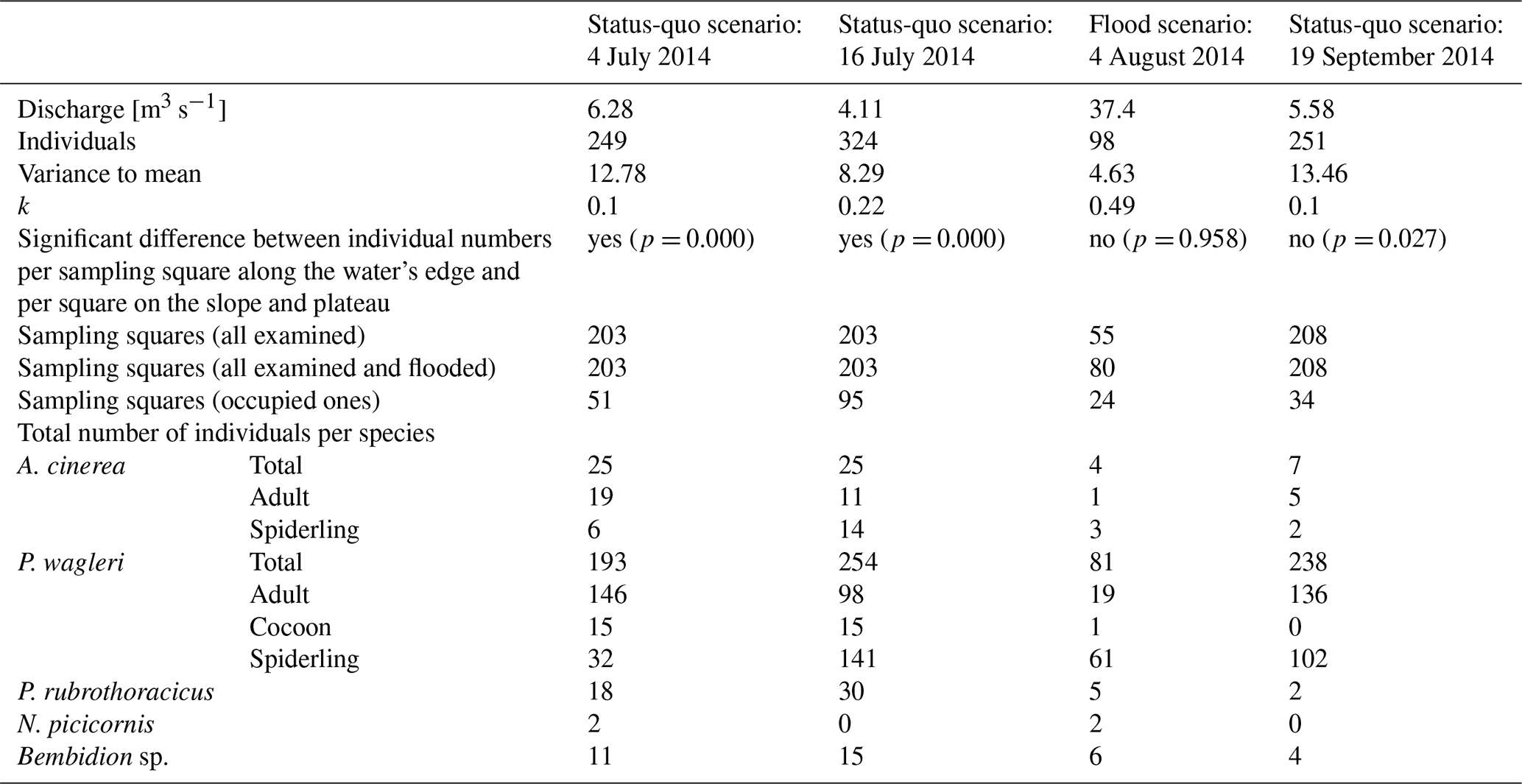
The study area on the gravel bank was partitioned into sampling squares measuring 5×5 m, resulting in a triangular study site approximately 105 m long and up to 75 m wide (Fig. 2). This configuration encompassed nearly half of the gravel bank. Each square was sampled once per sampling date, with a thorough manual examination conducted for arthropods. Stones were overturned, and finer substrates were sifted using fingers. Larger expanses of fine gravel and sand were carefully raked with a leaf rake. Individuals fleeing from the rake could be quickly spotted on the fine-grained substrate. Although this approach did not constitute a quantitative assessment, the effort for each sampling square was consistent and sufficiently high so that, in our estimation, an even greater effort or a fully quantitative assessment would not have led to fundamentally different results. The individuals of the studied species were counted, and their abundance per species within each square was documented. Sampling efforts were conducted on sunny days, ensuring there was no rain or fog, to ensure that many individuals were on the surface. The spatial distribution of the studied species was recorded three times under normal-flow conditions to establish a baseline or a “status-quo scenario”. Following a significant flood event, species distribution was recorded again to create a “flood scenario”. No surveys were conducted during smaller floods caused by frequent thunderstorms. A single major flood event occurred during the study period. The peak was at the end of July 2014, with a discharge of 152 m3 s−1 (detailed date information is provided in Table 1). A few days after the peak of the flood event, data for the flood scenario were recorded, at a discharge of 37.4 m3 s−1 (Gewässerkundlicher Dienst Bayern, 2025, and Table 1). At this time, the flood had exposed a large part of the gravel bank. An approximately 5 m wide, water-bearing flood channel, a remnant of the floodwaters that had not yet completely receded, separated the slope from the exposed gravel bank. The discharge values in Table 1 should be interpreted as guide values. The gauging station is located above the mouth of the Rißbach, and it is not known how much water from the Rißbach was discharged into the Isar during the flood event. The flood altered the dimension of the gravel bar, which made it necessary to adjust the number of sampling squares.
Table 1Number of individuals per sampling date and discharge scenario. Results are given for the variance-to-mean ratio, the clumping parameter k, and the Wilcoxon test.
2.2 Response of studied species to an artificial flood
To directly observe the response of the studied species to rising water levels, an artificially induced flood was created. On 25 July 2015, an artificial island composed of sand and gravel, approximately 1 m2 in area and about 25 cm in height, was constructed in a side channel approximately 3 m wide. A total of 37 individuals of the studied species were collected from adjacent gravel bars and islands and placed on the artificial island. Following a 1 h acclimatization period, the side channel was dammed until the artificial island was completely submerged, which occurred after about 15 min. The reaction of the individuals on the island was observed as the water level rose. Chronological observations were recorded to document whether individuals retreated from the rising water, fled the island, or exhibited other responses, thereby capturing the behavioural repertoire of the species under investigation.
2.3 Statistical analysis
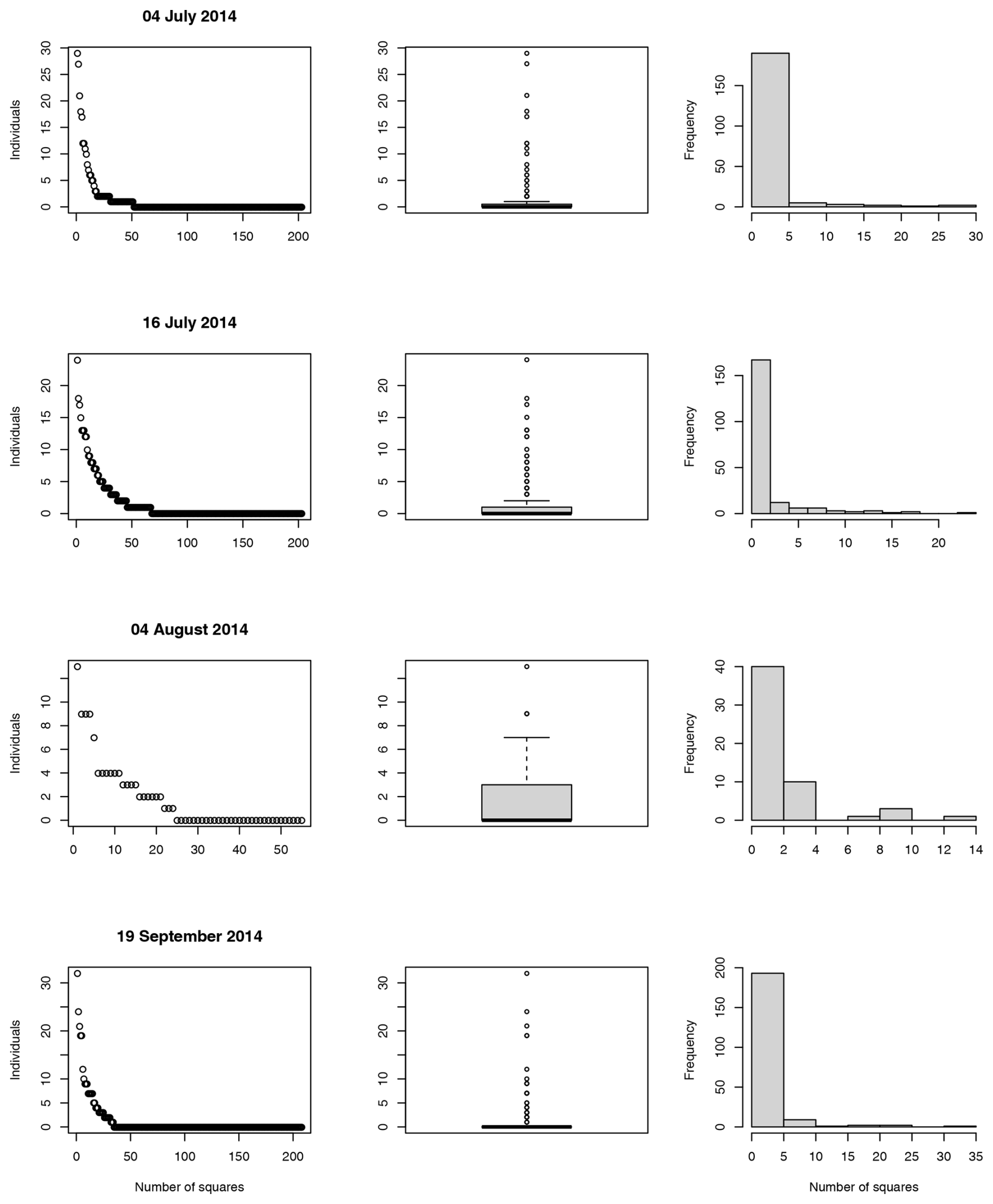
For a visual representation of the studied section of the gravel bar, maps were generated with QGIS version 3.16.13. Due to the constraints imposed by the nature reserve, aerial photos could not be captured on the sampling date due to the breeding season. Instead, maps were created using georeferenced aerial photographs taken the following November, with modifications made to reflect the sampling area and adjacent regions as documented on the sampling day. The data analysis was conducted using R version 4.1.2. In order to compare the data from the different sampling dates, the variance-to-mean ratio and the clumping parameter k were calculated for the total number of individuals observed. A variance-to-mean ratio of 1 shows a Poisson distribution, which indicates that the counts are randomly distributed. A variance significantly greater than the mean suggests that the counts are more variable than expected under a Poisson model and may indicate a clumped or aggregated distribution. The clumping parameter k assesses the degree of aggregation within the data, with k < 1 indicating high aggregation and k > 5 indicating randomness (Crawley, 2013). The Wilcoxon test was calculated to compare median values of individual counts of squares adjacent to the main water channel, with those in other squares located on the plateau and slope (Fig. 3), keeping in mind that rank transformation of counts can be unreliable when counts are low.
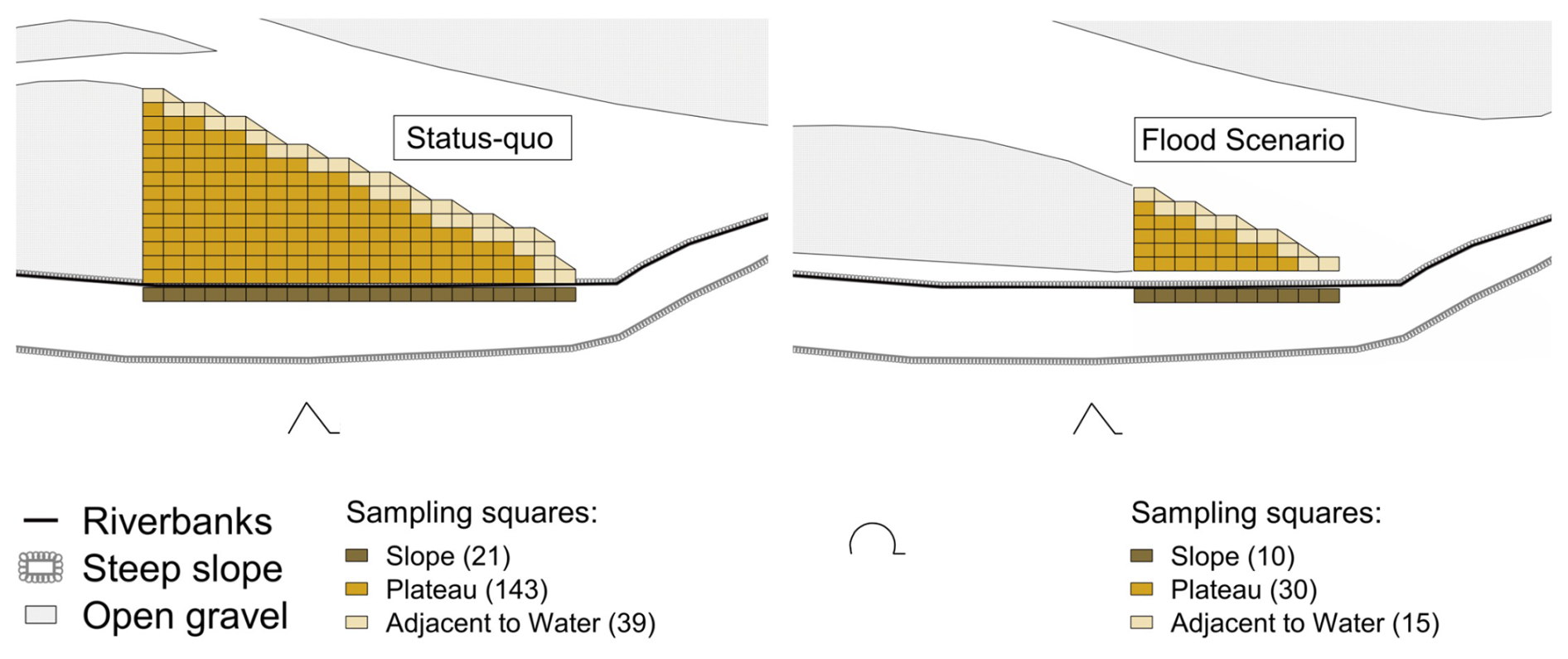
Figure 3Location and classification of the sampling squares adjacent to the main water channel (beige) and those located on the plateau (light brown) and at the slope (dark brown) for the status-quo (July) and flood scenario (August), with the number of sampling squares in brackets.
Unfortunately, the planned continuation of observations in 2015 could not be executed due to the complete erosion of the gravel bank during the winter. Consequently, further evaluations of the existing data were conducted through qualitative descriptions.
3.1 Change in species distribution pattern due to flooding
Altogether, 922 individuals of the studied species were counted across four different sampling dates (Table 1). Of these, P. wagleri accounted for 766 individuals, A. cinerea for 61 individuals, P. rubrothoracicus for 55 individuals, and the beetles for 40 individuals. Four of the beetles belonged to the species N. picicornis, while the rest belonged to the genus Bembidion sp. Even if there are slight differences between the individual distributions in the status quo before and after the flood, a clear differentiation from the distribution pattern during the flood can be seen (Fig. 4).
3.1.1 Pre-flood
The species distribution pattern (Fig. 5) is very similar for the individuals recorded on both pre-flood sampling dates, with a more frequent occurrence of individuals near the water and a sparser occurrence on the plateau. On both sampling dates, the number of individuals per square along the main water channel was significantly higher than in squares not adjacent to the water's edge (Wilcoxon rank-sum test p= 0.000–0.027). None of the examined species were found on the riverbank or the slope.
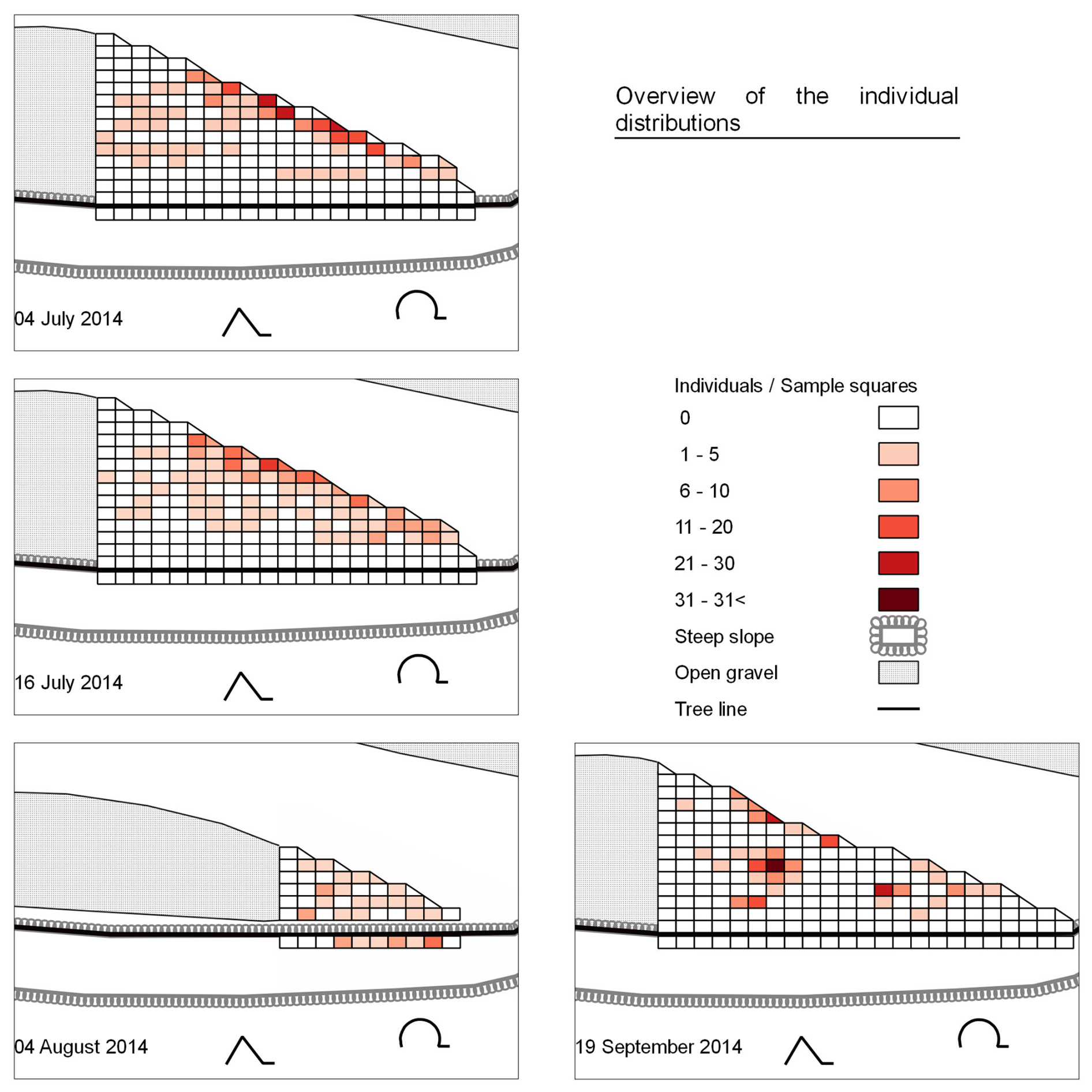
Figure 5Distribution of the studied species on different sampling dates. On 4 August 2014, data were collected 4 d after a flood that completely submerged the gravel bar. The extent of the gravel bank shown corresponds to the water level at the time of data collection.
3.1.2 Flood scenario
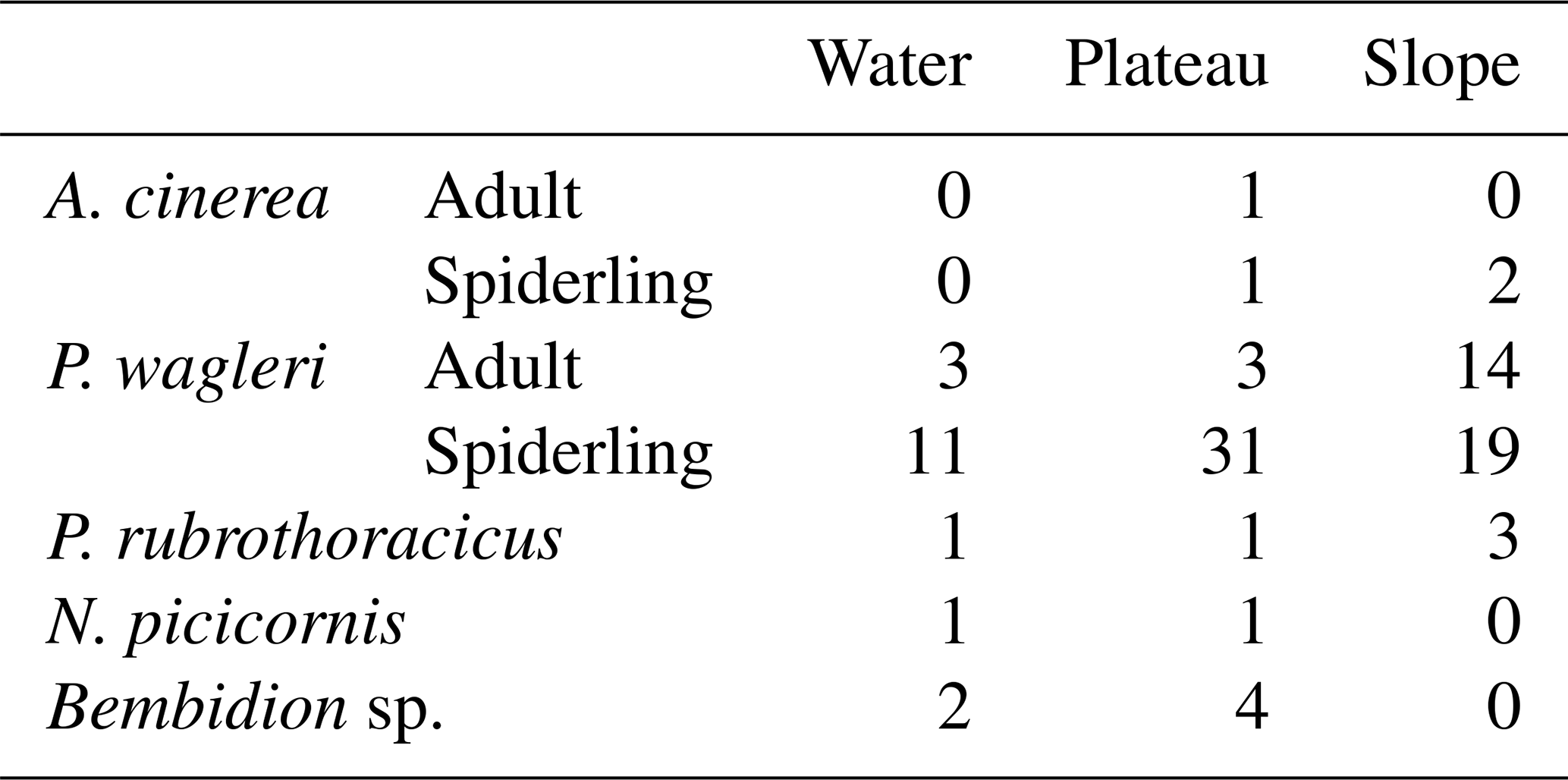
A different distribution pattern emerged during a flood. Of the individuals, 38.8 % were found on the slope, with an average of 3.8 individuals per sampling square (Table 2). Both spider species and the rove beetle were present (Table 3), but no ground beetles were found. The remaining individuals were scattered across the gravel bar, with an average of 1.3 individuals per sampling square. Two larger clusters were found around deadwood and smaller organic debris washed ashore. The Wilcoxon rank-sum test found no significant difference between the number of individuals found in squares next to the main water channel and those in the other squares on the plateau, riverbank, and slope (p= 0.958).
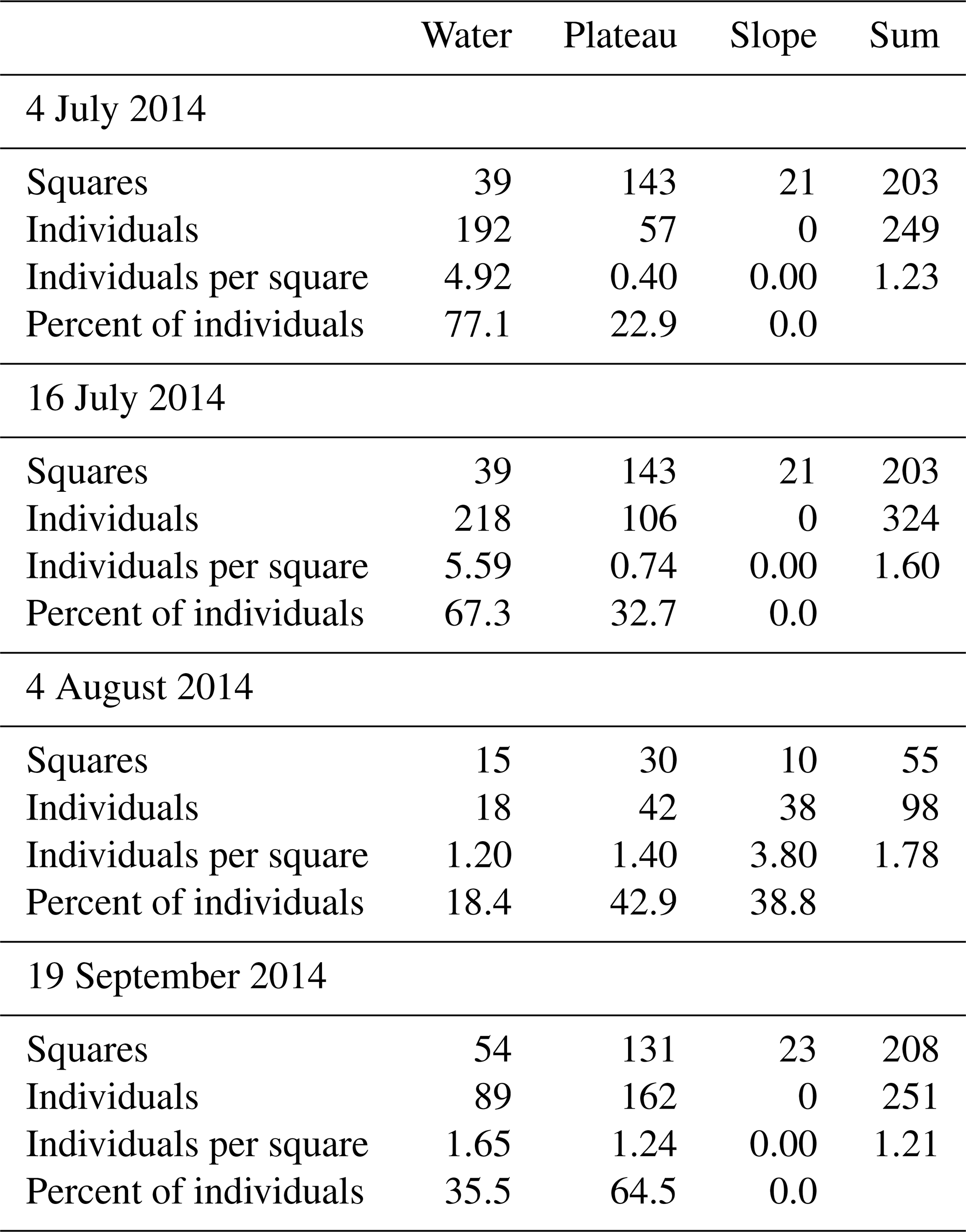
Table 3Number of individuals per species recorded during the flood scenario.
3.1.3 Post-flood
The distribution of individuals was again oriented towards the water's edge. On the plateau, some clusters of individuals were found on sampling squares with accumulations of organic matter. The differences between the number of individuals found in sampling squares next to the main water channel and those in other squares on the plateau and slope were not significant (p= 0.027) (compare also Fig. 5).
3.2 Response of the studied species to flooding
The individuals (Table 4) released on the artificial island quickly spread out and hid in the gravel. When they came to the surface of the water, they returned to the artificial island, drifted, or continued toward the opposite side of the water. When the rising water level drove the animals from their hiding places, this behaviour was repeated. As the area of the artificial island shrank, the animals increasingly ventured onto the water's surface. Shortly before the island was completely flooded, the animals no longer avoided the approaching waterline but ran directly across the water's surface to the opposite gravel bar upon contact with the water. In the end, only two P. wagleri individuals remained and moved across the water's surface toward a nearby gravel bar.
Table 4Number of individuals used in the behavioural experiment on rising water levels.
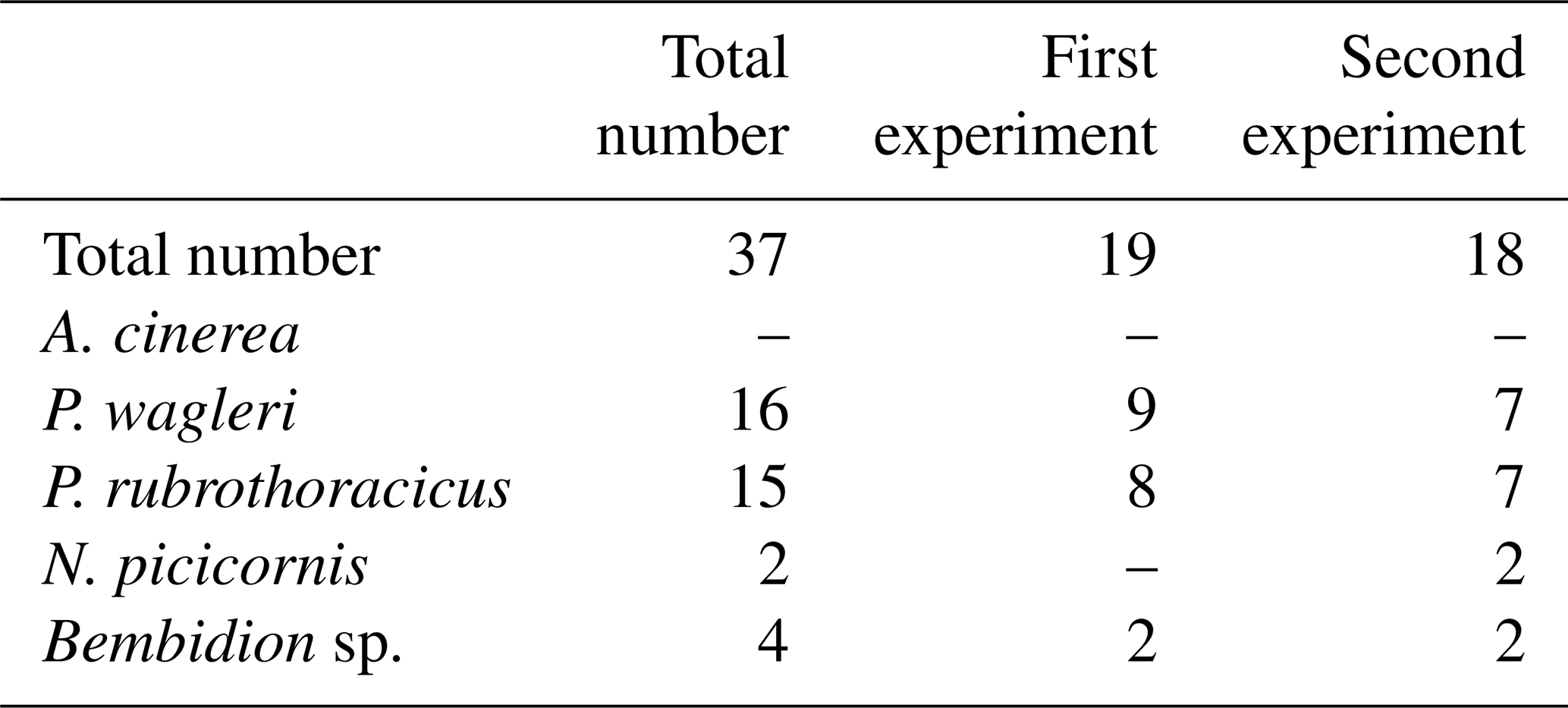
The Bembidion individuals also dodged the approaching waterline until the area became too small for them to retreat, at which point they paddled to a nearby gravel bar. One Bembidion had retreated to an elevated stone. When the rising water reached the Bembidion, it attempted to fly but failed. The two individuals of N. picicornis had already left the artificial island when the substrate became damp due to the rising water. They ran across the surface of the water to a nearby gravel bank.
To better understand the habitat requirement of the arthropod community on gravel bars, we examined the importance of elevated areas and riverbanks during flooding. The results indicate that elevated areas in the riverbed and riverbanks serve as refuge sites, actively visited by 39 % of the total number of collected individuals during a flood. Our study demonstrated that retreating to higher ground was a dominant response compared to fleeing across the water onto other gravel bars in reaction to rising water levels. Notably, the animals that retreated to the slope occupied a habitat they would typically have avoided, as noted by Wessels and Sundermann (2022), in the absence of flooding.
Some individuals might not have avoided the flood and instead survived in the inundated interstitial spaces, as evidenced by a single Bembidion specimen observed during this study, which is also supported by existing literature (Andersen, 1968; Manderbach, 1998; Kühnelt, 1943; Zulka, 1994a). It remains unclear whether the other individuals found on the plateau survived flooding in the interstitial spaces, migrated from the slope over the small water channel, or were washed up by rising water. Other described but unobserved behaviours of the species examined include diving to hide in the underwater interstitial, drifting on the water surface, and – for Carabidae – escaping through the air. However, it is unclear which criteria the animals use to determine their strategy in response to flooding.
4.1 Status-quo distribution
Gravel-bank-dwelling arthropods are typically found along the water's edge, a behaviour closely linked to food availability. Depending on the season, the primary food source for these arthropods consists of aquatic insects, which either drift from the water or emerge as adults (Briers et al., 2005; Hering and Plachter, 1997; Langhans and Tockner, 2014). During hatching events, the abundance of these insects creates a concentrated and readily accessible food supply along the water's edge. Consequently, arthropods are more likely to stay near the water's edge, where they can capitalize on this resource.
4.2 Flood-response-triggering factors
One criterion that may influence the nature of the response to floods is the current phase of the life cycle. For instance, the wolf spider Tricca lutetiana was observed to remain in its burrow at all times while it had a cocoon, without leaving under any circumstances that might lead to flooding (Dolejš et al., 2010). A similar situation was noted for a single female A. cinerea with a cocoon, which was located in an elevated, vegetated, and heavily silted area with a comparatively high abundance of ants, inside a burrow beneath a stone, with the entrance sealed by woven silk (Wessels and Sundermann, 2022). A female Arctosa maculata carrying a cocoon was also observed sitting in its submerged burrow, awaiting the end of the flood (Wessels and Sundermann, 2022). However, further literature on the wolf spider A. cinerea staying in its burrow during a flood does not provide information regarding the life phase of the individuals affected or whether they possessed a cocoon (Bellmann, 2010; Panait, 2005).
The nature of the response to floods can also be influenced by the current location of the affected individuals. For example, burrowed larvae of various Bembidion and Staphylinidae species survived flooding in the interstitial spaces, while individuals on the surface were carried away by the current (Andersen, 1968).
Another criterion that may determine the strategy employed is weather conditions. Bembidion individuals have been observed to fly from the water surface (Andersen, 1968), even at temperatures of around 16 °C during floods (Zulka, 1994a); however, they were unable to fly during winter (Manderbach, 1998) or on cool late-spring days with temperatures of around 10 °C (Wessels, unpublished data, 2014/15). On warm and sunny days, collecting individuals of the genus Bembidion proved challenging because they would fly away (unpublished data, 2014/15). This, along with the observation of a single Bembidion specimen that was unable to take off in the present study despite temperatures of around 22 °C, suggests that the ability to fly cannot be generalized across all individuals.
Temperature and light intensity significantly influenced the movement patterns of the studied Bembidion species. Individuals oriented toward the light at warm temperatures (15 °C and above) and toward dark silhouettes during colder temperatures (6 °C and below). This behaviour was independent of the experimental design (Andersen, 2006). This photosensitive reaction may explain behaviours such as the significantly decreased tendency of drifting individuals of the wolf spider Pardosa agricola to move ashore under sunny weather conditions (Lambeets et al., 2010). Similarly, individuals of P. wagleri were observed on warm summer days during data collection for the present study and other studies (Wessels and Sundermann, 2022), as they fled startled onto the water and remained some distance from the gravel bank, partially drifting away with the current. Overall, this response can lead to a migration to elevated areas during rainy weather to avoid a subsequent flooding. Conversely, on sunny days, such as the day the present study was conducted, this response increases the risk of individuals drifting, which may facilitate the recolonization of new gravel bars.
4.3 Population collapse and recovery induced by flooding
The literature describes that flooding leads to a collapse in surveyed arthropod populations (Hering et al., 2004; Manderbach and Plachter, 1997; Manderbach, 2001; Vanbergen et al., 2017). This phenomenon was also observed in our study, as fewer individuals were recorded following the flood. A detailed analysis of the carabid abundance at species level by Vanbergen et al. (2017) revealed that only wingless species were negatively affected by flooding. Due to high reproduction rates and strong dispersal abilities, the riparian arthropod communities typically recover within the same year (Bates et al., 2006; Hering et al.,2004; Kühnelt, 1943; Manderbach and Plachter, 1997; Manderbach, 2001). However, 1 month after the flood event, the number of individuals per square in our study approached the numbers before the flood event. However, it is unclear whether and, if so, what role animals that survived the flood in the interstitial played. Floods contribute to population recovery by resetting the successional stages (Larsen et al., 2019; Plachter, 1998), which helps preserve the system and provides a habitat for the gravel bar arthropod community. Additionally, flooding washes aquatic food onto riverine sediments (O'Callaghan et al., 2013; Tockner and Waringer, 1997). As a result, Hering et al. (2004) reported an increase in ground beetle species following a major flood. In the present study, the distribution of individuals across the sampling area on 19 September 2014 may indicate such an enrichment of food. Driftwood and organic matter washed ashore are known to harbour displaced arthropods (Tenzer, 2003; Zulka, 1994a), which may explain the clustering of individuals of the studied species that prey upon them.
4.4 Riverbanks and elevated areas as refuges during flooding
With more than 80 % of the riverbed being relocated during a single flood event (Van der Nat et al., 2003), the role of elevated areas within the riverbed and riverbanks in population recovery and resettlement is likely significant and should not be underestimated. The results of the present study, where 39 % of the total number of individuals were found on the riverbank during a flood, support this assumption. Indeed, elevated areas and riverbanks accessible to the riparian arthropods have been identified as important variables in several studies (Bell et al., 1999; Wessels and Sundermann, 2022). Abundance and, in some cases, species richness were the highest near elevated areas and riverbanks (Lambeets et al., 2009; Wessels and Sundermann, 2022), a trend that was also observed in organisms inhabiting interstitial spaces at a depth of 50 cm (Marmonier et al., 2019).
Although specific conditions required for riverbanks to function as flood refuges are not explicitly documented in the literature, it is likely that the stenotopic species of open gravel banks require conditions similar to those of their typical habitats. For example, the spider species A. cinerea and P. wagleri avoid areas with more than sparse vegetation (Framenau et al., 1996; Schatz et al., 2003; Wessels and Sundermann, 2022), possibly due to increased competition from generalist species or predators. Formicidae, with their efficient and aggressive foraging behaviour, are strong competitors, often outcompeting other arthropods for resources. Their dominance is further reinforced by their role as predators, as they directly hunt other gravel bank inhabitants, including arthropods (Hering, 1995; Wessels and Sundermann, 2022).
Riverbanks with these characteristics are found in semi-natural or natural river systems with lateral channel migration. In these systems, riverbanks, like the riverbed, are subject to regular erosion and deposition, creating the necessary refuges. This watercourse management should consider not only the riverbed but also the riverbanks and adjacent areas. This conclusion aligns with the recommendations of other authors (Rothenbücher and Schaefer, 2006; Ward et al., 2001) to protect species by safeguarding ecological processes at the landscape level.
All data used in this study are available in the Supplement, which provides code and the complete dataset supporting the results and conclusions of this article.
The supplement related to this article is available online at https://doi.org/10.5194/we-25-91-2025-supplement.
RW planned the campaign and analysed the data; RW and AS wrote the manuscript draft and reviewed and edited the paper.
The contact author has declared that neither of the authors has any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
The authors thank Harald Plachter, who gave the initial impetus for this project and was available for questions and discussions. The district government of Upper Bavaria is thanked for permission to collect the data. The authors also thank everyone who helped provide accommodation and support with data collection.
This paper was edited by Sergio Navarrete and reviewed by Arif Jan and one anonymous referee.
Andersen, J.: The effect of inundation and choice of hibernation sites of coleoptera living on river banks, Norsk Entomologisk Tidsskrift, 15, 115–133, 1968.
Andersen, J.: Mechanisms in the shift of a riparian ground beetle (carabidae) between reproduction and hibernation habitat, J. Insect Behav., 19, 545–558, 2006.
Bates, A. J., Sadler, J. P., and Fowles, A. P.: Condition-dependent dispersal of a patchily distributed riparian ground beetle in response to disturbance, Oecologia, 150, 50–60, 2006.
Bell, D., Petts, G. E., and Sadler, J. P.: The distribution of spiders in the wooded riparian zone of three rivers in Western Europe, Regul. Riv., 15, 141–158, 1999.
Bellmann, H.: Der Kosmos Spinnenführer, Franckh-Kosmos Verlags-GmbH & Co. KG, Stuttgart, ISBN 978-3-440-10114-8, 2010.
Bonn, A.: Flight activity of carabid beetles on a river margin in relation to fluctuating water levels, in Natural History and Applied Ecology of Carabid Beetles, edited by: Brandmayr, P., Lövry, G. L., Zetto Brandmayr, T., Casale, A., and Vigna Taglianti, A., Pensoft Publishers, Moscow, 147–160, https://archive.org/details/bonn-2000/mode/2up (last access: 2 May 2025), 2000.
Briers, R. A., Cariss, H. M., Geoghegan, R., and Gee, J. H. R.: The lateral extent of the subsidy from an upland stream to riparian lycosid spiders, Ecography, 28, 165–170, 2005.
Campolo, M., Andreussi, P., and Soldati, A.: River flood forecasting with a neural network model, Water Resour. Res., 35, 1191–1197, 1999.
Crawley, M. J.: The R book, second edition, Wiley & Sons Ltd, ISBN 978-0-470-97392-9, 2013.
Desender, K.: Ecomorphological adaptations of riparian carabid beetles, Comptes Rendus du Symposium “Invertebres de Belgique”, Royal Institute of Natural Sciences, Brussels, 309–314, https://biblio.naturalsciences.be/rbins-publications/miscellanous/verhandelingen-van-het-symposium-invertebraten-van-belgie-comptes-rendus-du-symposium-invertebres-de-belgique-proceedings-of-the-symposium-invertebrates-of-belgium/wouters-1989-proceedings-of-the-symposium-invertebrates-of-belgium-309-314.pdf (last access: 22 April 2025), 1989.
Dolejš, P., Kubcová, L., and Buchar, J.: Courtship, mating, and cocoon maintenance of Tricca lutetiana (Araneae: Lycosidae), J. Arachnol., 38, 504–510, 2010.
Eyre, M. D. and Lott, D. A.: Invertebrates of exposed riverine sediments, Techreport W11, Entomological Monitoring Dervices, https://assets.publishing.service.gov.uk/media/5a7c5a3840f0b660183b6ca3/str-w11-e-e.pdf (last access: 22 April 2025), 1997.
Ferguson, R. and Ashworth, P.: Spatial patterns of bedload transport and channel change in braided and near-braided rivers, in: Dynamics of Gravel-Bed Rivers, edited by: Billi, P., Hey, R. D., Thorne, C. R., and Tacconi, P., John Wiley and Sons Ltd, 477–496, ISBN 0-471-92976-X, 1992.
Framenau, V., Dieterich, M., Reich, M., and Plachter, H.: Life cycle, habitat selection and home ranges of Arctosa cinerea (Fabricius, 1777) (Araneae: Lycosidae) in a braided section of the Upper Isar (Germany, Bavaria), Rev. Suisse Zool., 223–234, 1996.
Gewässerkundlicher Dienst Bayern: Gesamtzeitraum Rißbachdüker/Isar, https://www.gkd.bayern.de/de/fluesse/abfluss/isar/rissbachdueker-16001303/gesamtzeitraum/tabelle#top, last access: 18 January 2025.
Hebets, E. A. and Chapman, R. F.: Surviving the flood: plastron respiration in the non-tracheate arthropod Phrynus marginemaculatus (Amblypygi: Arachnida), J. Insect Physiol., 46, 13–19, 2000.
Hering, D.: Nahrung and Nahrungskonkurrenz von Laufkäfern und Ameisen in einer nordalpinen Wildflussaue, Archiv für Hydrobiologie, Supplementband, Large rivers, Vol. 9, 439–453, https://doi.org/10.1127/lr/9/1996/439, 1995.
Hering, D. and Plachter, H.: Riparian ground beetles (Coeloptera, Carabidae) preying on aquatic invertebrates: a feeding strategy in alpine floodplains, Oecologia, 111, 261–270, 1997.
Hering, D., Gerhard, M., Manderbach, R., and Reich, M.: Impact of a 100-year flood on vegetation, benthic invertebrates, riparian fauna and large woody debris standing stock in an alpine floodplain, River Res. Appl., 20, 445–457, 2004.
Junk, W.: The flood pulse concept of large rivers: learning from the tropics, Large Rivers, 11, 261–280, https://doi.org/10.1127/lr/11/1999/261, 1999.
Kühnelt, W.: Die litorale Landtierwelt ostalpiner Gewässer, Internationale Revue der gesamten Hydrobiologie und Hydrographie, 43, 430–457, 1943.
Lambeets, K. and Bonte, D.: Between-population variation in homeward orientation behaviour in two riparian wolf spiders, Behav. Process., 82, 62–66, 2009.
Lambeets, K., Vandegehuchte, M. L., Maelfait, J.-P., and Bonte, D., Integrating environmental conditions and functional life-history traits for riparian arthropod conservation planning, Biol. Conserv., 142, 625–637, 2009.
Lambeets, K., Van Ranst, J., and Bonte, D.: Is movement behavior of riparian wolf spiders guided by external or internal information, J. Arachnol., 38, 313–318, https://doi.org/10.1636/P09-52.1, 2010.
Langhans, S. and Tockner, K.: Edge effects are important in supporting beetle biodiversity in a gravel-bed river floodplain, PlOS One, 9, e114415, https://doi.org/10.1371/journal.pone.0114415, 2014.
Larsen, S., Karaus, U., Claret, C., Sporka, F., Hamerlik, L., and Tockner, K.: Flooding and hydrologic connectivity modulate community assembly in a dynamic river-floodplain ecosystem, PLOS One, 14, e0213227, https://doi.org/10.1371/journal.pone.0213227, 2019.
Lehmann, H.: Ökologische Untersuchungen über die Carabidenfauna des Rheinufers in der Umgebung von Köln, Zeitschrift für Morphologie und Ökologie der Tiere, 55, 597–630, 1965.
Leopold, L. B. and Wolman, M. G.: River channel patterns: braided, meandering, and straight, US Government Printing Office, https://doi.org/10.3133/pp282B, 1957.
Manderbach, R.: Lebensstrategien und Verbreitung terrestrischer Arthropoden in schotterreichen Flußauen der Nordalpen, Görich und Weiershäuser, ISBN 9783897032446, 3897032449, 1998.
Manderbach, R.: Der Stellenwert des Lebenszyklus für das Überleben der uferbewohnenden Wolfspinnenarten Pardosa wagleri (Hahn, 1822) und Pirata knorri (Scopoll, 1763), Arachnol. Mitt., 21, Basel, 1–13, https://doi.org/10.5431/aramit2101, 2001.
Manderbach, R.: Laufkäfergemeinschaften am Ufer schotterreicher Fließgewässer der Nordalpen, Angewandte Carabidologie, 4, 33–40, 2002.
Manderbach, R. and Plachter, H.: Life strategy of the carabid beetle Nebria picicornis (Fabr. 1801) (Coleoptera, Carabidae) on river banks, Beiträge der Gesellschaft für Ökologie, 3, 17–27, 1997.
Marmonier, P., Olivier, M.-J., Des Châtelliers, M. C., Paran, F., Graillot, D., Winiarski, T., Konecny-Dupré, L., Navel, S., and Cadilhac, L.: Does spatial heterogeneity of hyporheic fauna vary similarly with natural and artificial changes in braided river width?, Sci. Total Environ., 689, 57–69, 2019.
O'Callaghan, M. J., Hannah, D. M., Boomer, I., Williams, M., and Sadler, J. P.: Responses to river inundation pressures control prey selection of riparian beetles, PLOS One, 8, e61866, https://doi.org/10.1371/journal.pone.0061866, 2013.
Panait, V.: The influence of the soil parameters upon the soil's mesofauna in function of the environment conditions within Mugrighiol-Dunavat dammed and drainage area as part of the Danube Delta Biosphere Reserve, Analele Ştiinţifice ale Universităţii “Al.I. Cuza” Iaşi, s. Biologie animală, http://cercetare.bio.uaic.ro/publicatii/anale_zoologie/issue/2005/16-2005.pdf (last access: 22 April 2025), 2005.
Papi, F.: Astronomische Orientierung bei der Wolfspinne Arctosa perita (latr.), Z. Vergl. Physiol., 37, 230–233, 1955.
Perevozkin, V., Lukyantsev, S., and Gordeev, M.: Comparative analysis of foraging behavior in aquatic and semiaquatic spiders of the genera Argyroneta, Dolomedes, Pirata, and Pardosa, Russ. J. Ecol., 35, 103–109, 2004.
Plachter, H.: Die Fauna der Kies-und Schotterbänke dealpiner Flüsse und Empfehlungen für ihren Schutz, Akad. für Naturschutz und Landschaftspflege, vol. 10, 119–147, https://www.zobodat.at/pdf/Ber-Bayer-Akad-f-Natursch-u-Landschaftspfl_10_1986_0119-0147.pdf (last access: 22 April 2025), 1986.
Plachter, H.: Die Auen alpiner Wildflüsse als Modelle Störungsgeprägter ökologischer Systeme, Schriftenreihe für Landschaftspflege und Naturschutz, 56, 21–66, 1998.
Rothenbücher, J. and Schaefer, M.: Submersion tolerance in floodplain arthropod communities, Basic Appl. Ecol., 7, 398–408, 2006.
Schatz, I., Steinberger, K.-H., and Kopf, T.: Auswirkungen des Schwellbetriebes auf uferbewohnende Arthropoden (Aranei; Insecta: Coleoptera: Carabidae, Staphylinidae) am Inn im Vergleich zum Lech (Tirol, Österreich), Amt der Tiroler Landesregierung, Abteilung Umweltschutz – 12, 202–231, https://www.zobodat.at/pdf/Natur-in-Tirol_12_0202-0231.pdf (last access: 22 April 2025),, 2003.
Tenzer, C.: Ausbreitung terrestrischer Wirbelloser durch Fließgewässer, PhD thesis, Philipps-University Marburg, https://archiv.ub.uni-marburg.de/diss/z2004/0242/pdf/dct.pdf (last access: 22 April 2025), 2003.
Tockner, K. and Waringer, J.: Measuring drift during a receding flood: Results from an austrian mountain brook (Ritrodat-Lunz), Internationale Revue der gesamten Hydrobiologie und Hydrographie, 82, 1–13, 1997.
Tockner, K., Paetzold, A., Karaus, U., Claret, C., and Zettel, J.: Ecology of braided rivers, Special Publication, Int. As. Sed., 36, 339, https://doi.org/10.1002/9781444304374.ch17, 2006.
Vanbergen, A. J., Woodcock, B. A., Gray, A., Andrews, C., Ives, S., Kjeldsen, T. R., Laize, C. L., Chapman, D. S., Butler, A., and O'Hare, M. T.: Dispersal capacity shapes responses of river island invertebrate assemblages to vegetation structure, island area, and flooding, Insect Conserv. Diver., 10, 341–353, 2017.
Van der Nat, D., Tockner, K., Edwards, P. J., Ward, J., and Gurnell, A. M.: Habitat change in braided flood plains (Tagliamento, NE-Italy), Freshwater Biol., 48, 1799–1812, 2003.
Ward, J. and Tockner, K.: Biodiversity: towards a unifying theme for river ecology, Freshwater Biol., 46, 807–819, 2001.
Ward, J. V., Tockner, K., Edwards, P. J., Kollmann, J., Bretschko, G., Gurnell, A. M., Petts, G. E., and Rossaro, B.: A reference river system for the alps: the fiume tagliamento, River Res. Appl., 15, 63–75, 1999.
Ward, J. V., Tockner, K., Uehlinger, U., and Malard, F.: Understanding natural patterns and processes in river corridors as the basis for effective river restoration, Regul. Rivers, 17, 311–323, 2001.
Weigmann, G. and Wohlgemuth-von Reiche, D.: Vergleichende Betrachtungen zu den Überlebensstrategien von Bodentieren im Überflutungsbereich von Tieflandauen, Limnol. Akt., 9, 229–240, 1999.
Wessels, R. and Sundermann, A.: Habitat requirements of riparian arthropods on gravel bars: Implications for conservation and management of braided river floodplains, PLOS One, 17, e0274977, https://doi.org/10.1371/journal.pone.0274977, 2022.
Wohl, E.: Rivers in the Landscape, John Wiley & Sons, ISBN 9781118414897, 2014.
Zulka, K. P.: Carabids in a central european floodplain: species distribution and survival during inundations, Carabid beetles: ecology and evolution, Springer, 399–405, https://doi.org/10.1007/978-94-017-0968-2_61, 1994a.
Zulka, K. P.: Natürliche Hochwasserdynamik als Voraussetzung für das Vorkommen seltener Laufkäferarten (Coleoptera, Carabidae), Wiss. Mitt. Niederösterr. Landesmuseum, 8, 203–215, 1994b.