the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 05 Feb 2024
| 05 Feb 2024
Bird color and taxonomic diversity are negatively related to human disturbance in urban parks
Lucas M. Leveau
Juan Kopp
Recently, studies have shown that highly urbanized areas are numerically dominated by birds with grey plumage, probably favoring camouflage with impervious grey surfaces. However, patterns of bird color diversity in urban green areas remain unexplored. In urban parks, sites surrounded by highly urbanized areas may be dominated by grey bird species, whereas urban parks with pedestrians that feed birds may favor commensal birds which also have grey plumage. Thus, we explore the relationship between environmental variables and bird taxonomic and color diversity in urban parks. Bird surveys were conducted twice by fixed points in urban parks of six cities in central Argentina. Bird color diversity was assessed by characterizing bird plumage coloration in different parts of the bird body and the presence of plumage sexual dimorphism, polymorphism, and iridescence. Then, color richness and color diversity (abundance-based) were calculated with species mean pairwise distance. Null models were used to quantify richness-corrected color diversity. Bird species richness and diversity increased with green-area size, and bird diversity decreased with increasing pedestrian traffic. Color richness decreased with increasing car traffic, whereas color diversity was positively related to green-area size and negatively related to car and pedestrian traffic. Richness-corrected color diversity related negatively to car and pedestrian traffic. The abundance of grey birds increased in parks near the urban centers and related positively to pedestrian and car traffic. The abundance of yellow birds increased in parks near rural areas or small cities. Color diversity increased with green-area size, but richness-corrected color diversity ceased to be related to area size. Therefore, the increase in color diversity with area size was mediated by increases in species richness. Color diversity was clustered in parks with the highest human disturbance, favoring the abundance of grey species which probably took advantage of food discarded by humans.
- Article
(2053 KB) - Full-text XML
-
Supplement
(422 KB) - BibTeX
- EndNote




The theory of island biogeography (MacArthur and Wilson, 1967) predicts that bird species richness increases on those largest islands near the continental pool of species. The greater area size promotes lower species extinction, whereas the nearness to the continental species pool promotes greater species colonization. These predictions have also been tested in fragments of terrestrial habitats or habitat islands surrounded by hostile habitats, such as forest fragments and urban green areas (Haila, 2002; Matthews et al., 2016). However, other confounding factors correlated with area size such as habitat diversity can also play a role in molding species richness in habitat islands (Simberloff, 1976; Ricklefs and Lovette, 1999). Habitat diversity, measured as the proportion of vegetation strata or different habitat types in a site, may enhance species diversity by providing different resources to birds (MacArthur and MacArthur, 1961; Tews et al., 2004).
Urban parks are considered hot spots of bird diversity in cities (Nielsen et al., 2014). Due to urban parks being heavily designed and managed, their habitat diversity may be independent of their area size. Therefore, urban parks are excellent places to test the relative roles of area size, isolation, and habitat diversity molding bird diversity (Leveau and Leveau, 2016; Yang et al., 2020). Moreover, human disturbances such as noise or pedestrian traffic may also negatively impact species diversity and influence species composition (Fernández-Juricic, 2002; González-Oreja et al., 2012; Zhou and Chu, 2012). In addition, urban green areas are threatened by urban expansion within cities or urban densification (Colding et al., 2020). Therefore, understanding of the relationship between green-area size, isolation, habitat diversity, and bird diversity is fundamental to predicting the effects of urban densification in cities.
Bird diversity in urban parks has been explored through different facets, such as taxonomic, functional, or phylogenetic diversities (Morelli et al., 2017; Curzel et al., 2021; Nava-Díaz et al., 2022; Melo et al., 2022; Mao et al., 2023). Due to the functional and phylogenetic diversities being generally positively correlated with species richness (but see Morelli et al., 2016, for birds and Lopez et al., 2018, for vascular plants), null models are used to analyze the departures of the functional and phylogenetic diversity with respect to chance (Petchey et al., 2007). Therefore, positive values compared to the null model indicate overdispersion of functional traits or phylogenetic relatedness, whereas negative values indicate clustering of functional traits or phylogenetic relatedness (Webb et al., 2002; De Bello et al., 2021).
Studies that have analyzed departures from null models of functional or phylogenetic traits in urban green areas are scarce (Zhang et al., 2018; Liu et al., 2019; Nava-Díaz et al., 2022). In general, these studies have found that increased urban park size is related to clustered functional and phylogenetic traits. Therefore, the larger urban green areas are inhabited by species with similar functional traits and that are phylogenetically related. This pattern has been found in a recent global meta-analysis (Matthews et al., 2023), and its authors proposed that the number of guilds increased with area size at a slower rate than the number of species, thus promoting functional redundancy. The functional diversity considered in these studies is related to the variety of food resources species obtain. Other traits such as bird plumage color (bird color hereafter) may have a role in filtering species according to area size, isolation, or habitat diversity. However, the analysis of bird plumage diversity has not yet been analyzed in urban green areas.
Bird color diversity has recently been analyzed along urbanization gradients, and sites dominated by buildings were clustered compared to null models, being dominated by bird species with grey or dark colors (Leveau, 2019; Turak et al., 2022). Grey plumage may enhance bird camouflage with impervious grey surfaces (Leveau, 2019). Therefore, the match of color plumage and habitat surfaces to avoid predation could be a significant factor in molding species' presence in anthropogenic environments (Bailey, 1978; Endler, 1993; Delhey and Peters, 2017; Leveau, 2021; Leveau and Ibañez, 2022). Birds in urban environments face predation risk mainly by avoiding humans, cats, and dogs (Møller and Ibáñez-Álamo, 2012; Morelli et al., 2023). In addition, plumage colorfulness could be related to bird intraspecific communication in closed urban environments, such as wooded urban parks (Butcher and Rohwer, 1989; Cooney et al., 2022). On the other hand, bird plumage color in urban parks may be influenced by the surrounding urban landscape, favoring the dominance of grey species in parks surrounded by high levels of urbanization. Moreover, the presence of pedestrians who feed birds may also favor the presence of commensal bird species, which usually have grey plumage color, such as doves or house sparrows (Passer domesticus) (Hambuckers et al., 2023; Swartz et al., 2023).
On the other hand, the pattern of bird color diversity and composition in urban areas is useful for understanding the loss of cultural ecosystem services provided by birds (Goodness et al., 2016; Valente-Neto et al., 2021). Bird colors have been important for human perception in urban areas (Belaire et al., 2015; Andrade et al., 2022). For example, yellow, orange, and blue plumage colors have been appreciated by humans (Lišková and Frynta, 2013; Frynta et al., 2010). Although the information about bird color composition and urban green areas is scarce, published studies have found that green areas surrounded by buildings or noise are dominated by grey species (Hambuckers et al., 2023; Swartz et al., 2023).
The objective of this study was to analyze the variation in taxonomic and color diversity in urban parks of six cities in central Argentina. Regarding taxonomic diversity, we expect greater bird diversity in the larger and less isolated urban parks. Moreover, we expect a positive relationship between species diversity and habitat diversity. Regarding bird color diversity, we expect that urban parks surrounded by buildings will be numerically dominated by bird species adapted to impervious areas and human presence, those with grey plumage and an omnivorous diet to exploit discarded human food. Urban areas near to city fringes and surrounded by less impervious cover may have a higher diversity of bird colors. Based on previous studies (Nava-Díaz et al., 2022; Matthews et al., 2023), we expect a clustering of plumage colors in the larger urban parks.
2.1 Study area
The study was conducted in the pampa grassland of central Argentina (Fig. 1). The climate is subtropical temperate with cold winters and hot and wet summers. The original vegetation comprises hygrophyte and halophyte meadows associated with edaphic–topographic conditions, where we can find species such as Piptochaetium stipoides, Piptochaetium montevidense, Solanum glaucophyllum, Sporobolus pyramidatus, and Sporobolus montevidensis among others (Oyarzabal et al., 2018). In addition, Celtis ehrenbergiana forests can be found associated with calcareous shell deposits parallel to the Atlantic coast (Parodi, 1940). Today the region is heavily impacted by agriculture, cattle raising, and tree plantations.
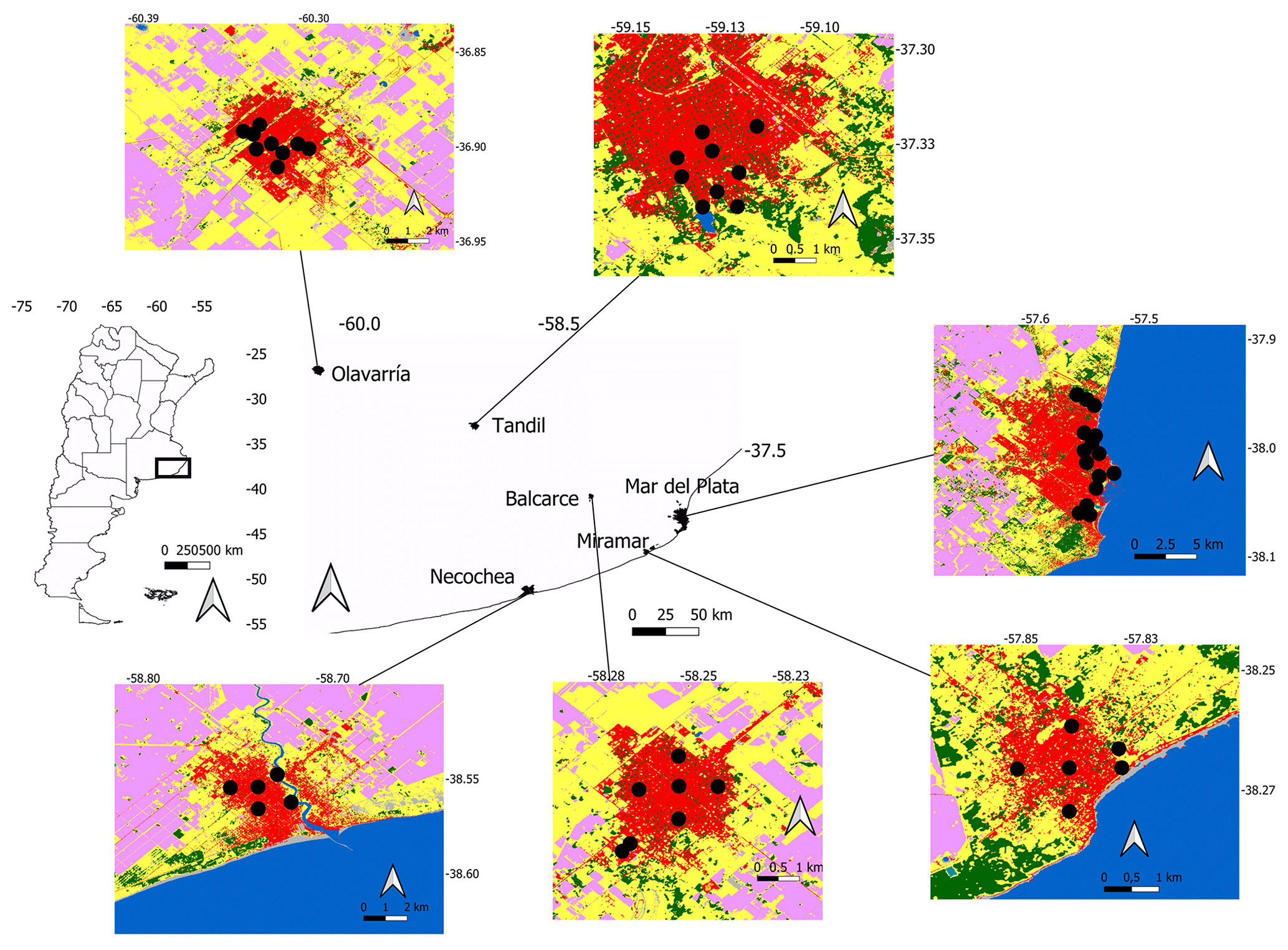
Figure 1Location of urban parks in six cities of central Argentina. The black dots on the city maps indicate the parks that were studied. Red: urban area; green: forests; yellow: lawn or grasslands; pink: cultivated area; blue: lakes, rivers, or sea. Images were obtained from WorldCover 2021 v200 (https://esa-worldcover.org/en, last access: 29 October 2021).
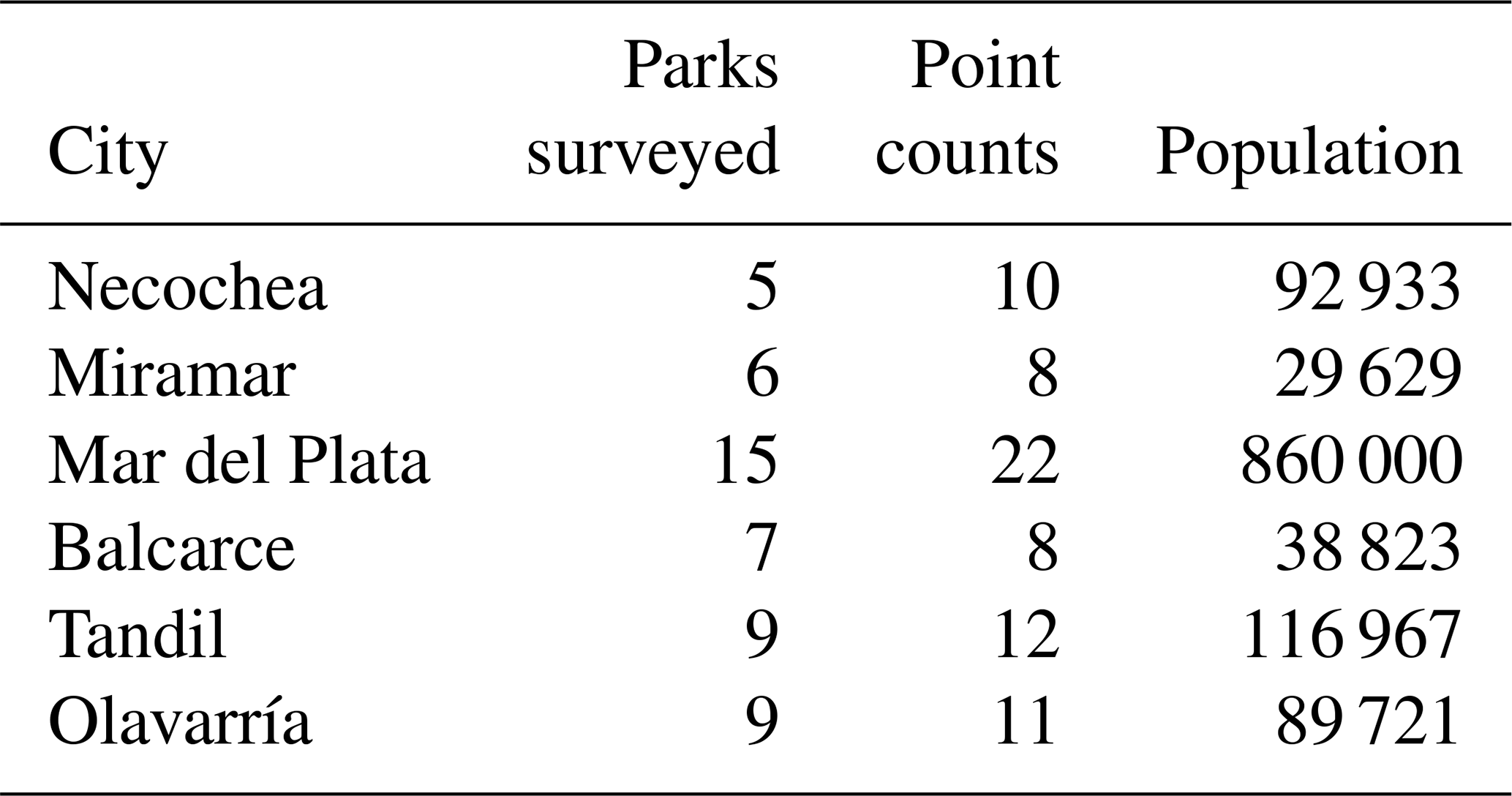
A total of 51 urban parks in six cities were surveyed. Green areas were urban parks composed generally of managed exotic vegetation and asphalted sidewalks. Human population size ranged between 860 000 inhabitants (Mar del Plata) and 29 629 inhabitants (Miramar), with a population increase between 4 % (Necochea) and 22 % (Mar del Plata) (National Institute of Statistics and Censuses, period 2000–2010) (Table 1). Population increases were related to the process of urban expansion over rural areas and urban densification (Merlotto et al., 2012; Leveau, 2014; Linares et al., 2022). Cities were founded between 1823 (Tandil) and 1888 (Mirarmar).
2.2 Bird surveys
Bird counts were performed during the first 4 h of the morning by an observer with more than 15 years of experience. Two visits were made: one during autumn (July) and one during spring (October–November) 2018. The surveys were carried out at fixed points with a radius of 50 m for 5 min, in parks separated by at least 200 m from each other. In parks smaller than 2 ha, a single point was established in the center of the park. In larger parks, two or more points were located that were separated by at least 200 m. Only birds that used the park, either eating or perching, were counted, avoiding birds that flew high above.
2.3 Environmental variables
At each sampling point, four perpendicular photographs were taken according to the four cardinal points to make an estimate of the habitat structure. For each of the photographs, the percentage cover of grass, trees, shrubs, asphalted sidewalks, buildings, and bare soil was estimated visually. The values of the four photographs were averaged for each point. With these cover values, habitat diversity was quantified and measured with the Shannon index (Shannon, 1948) based on the proportion data of each substrate. During the counts, the number of people at each point and the number of cars passing on the street closest to the point were also quantified. Finally, the size of each park was measured with Google Earth Pro.
At the landscape scale, the minimum distance from each park to rural areas, the minimum distance to other green areas equal to or greater than 1 ha, and the minimum distance to the city center were measured using Google Earth Pro. Rural areas were defined as sites outside city polygons (Fig. 1) with building densities lower than 2.5 buildings ha−1 (Marzluff et al., 2001). The center of each city was considered the most urbanized area of the city and was characterized by an urban park surrounded by the administrative and the commercial center. The number of inhabitants of each city was also obtained based on data from the last national census. The number of inhabitants is an indicator of the size of the city which would act on the dispersal of birds. The larger the size, the greater the isolation of non-urban bird populations is expected to be.
2.4 Bird plumage color
Bird plumage color (hereafter bird color) was obtained through plates from the Handbook of the Birds of the World Alive (HBWA, https://www.hbw.com, last access: 7 May 2020) and from the Aves Argentinas free mobile phone application, both consulted during January 2019. Color types were taken from 12 patches of the bird body (see Leveau, 2019, and Table S1 in the Supplement): (1) crown, (2) head, (3) cheek, (4) mantle, (5) primaries, (6) wing coverts, (7) back, (8) rump, (9) tail, (10) belly, (11) breast, and (12) throat. When the rump color description was not available, we searched for photographs on Google where the rump was visible. Color characterization was obtained from color descriptions in HBWA, the Aves Argentinas phone application, and personal consideration from one of us (Lucas M. Leveau). We created binary categories for body patches that had two colors in which the first color was the dominant (see Leveau, 2019). Other plumage characteristics were the presence of sexual plumage dimorphism, polymorphism, and iridescence (Table S1).
2.5 Taxonomic and color diversity
Taxonomic diversity was estimated by species richness and diversity. Species richness was the accumulated species in each green area during the two visits. Species diversity was the Shannon index calculated with accumulated bird abundances during the two visits for each green area.
Color richness was calculated with the mean pairwise distance between species pairs in the community matrix. A Gower distance was calculated for the matrix of species colors, and then a presence–absence community matrix was used. Color diversity was calculated with the mean pairwise distance weighted by species abundances. Color richness and diversity were calculated with the function mpd in picante (Kembel et al., 2010; R Core Team, 2019). Because color richness and diversity may be influenced by species richness values in each sampling unit, standardized effect sizes of mean pairwise distances for richness (SES richness) and diversity (SES diversity) were calculated using the null model independent swap through 999 randomizations with the function ses.mpd in picante (Kembel et al., 2010). Positive SES values indicate that bird color richness or diversity is higher than expected by chance, and negative SES values indicate that color richness or diversity is lower than expected by chance.
2.6 Color composition
The relationship between color composition and environmental variables was assessed through fourth-corner analysis (Brown et al., 2014). This analysis allows fitting a predictive model for species abundance as a function of environmental variables, species traits, and their interaction. Each species was classified according to the predominant color in their plumage patches and the presence of sexual dimorphism, polymorphism, and iridescence (Table S2 in the Supplement). This matrix was compared to the species abundances by site matrix and to the environmental variables by site matrix with the function traitglm of the mvabund package, and a Gaussian structure of errors was used (Wang et al., 2018).
2.7 Statistical analysis
The relationship between taxonomic and color diversity and environmental variables was assessed through generalized linear models. For species richness, a Poisson error structure was used. For the rest of the response variables, a Gaussian error structure was used. Multicollinearity was assessed through Pearson correlations between environmental variables, considering a correlation >0.70 to be important (Vatcheva et al., 2016). A significant correlation was found between human population and distance to rural areas (r>0.70); therefore we used only distance to rural areas in models. A backward model selection was used from the full model using the ANOVA function. Final models were compared with null models using a likelihood ratio test (LRT test) (P<0.05). The r2 of significant models was obtained with the function rsq of the package rsq (Zhang, 2021). All diagnostic analyses were carried out with the DHARMa package (Hartig, 2021). Due to parks being nested within cities, we explored significant (P<0.05) spatial autocorrelations of models residuals with SAM software (Rangel et al., 2010) and found no significant patterns of spatial autocorrelation.
Bird species richness and taxonomic diversity increased with increasing green-area size (richness: LRT = 27.82, P<0.001, r2=0.44; diversity: LRT = 2.57, P<0.001, r2=0.45; Table 2a, b; Fig. 2a, b). Moreover, bird taxonomic diversity was negatively related to pedestrian traffic (Table 2b; Fig. 2c).
Table 2Final generalized linear models between taxonomic and color diversity and environmental variables in urban parks of six cities in central Argentina.
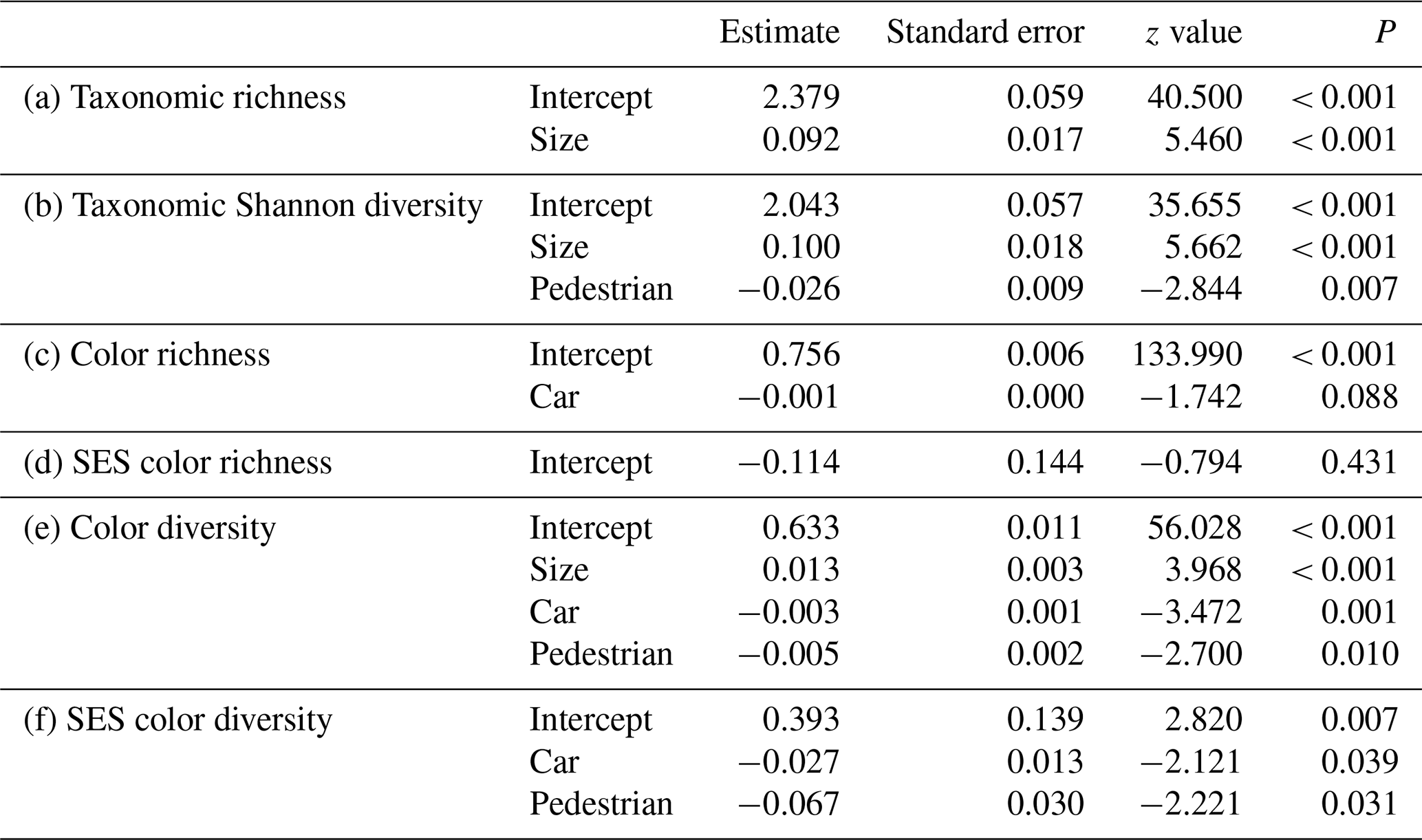
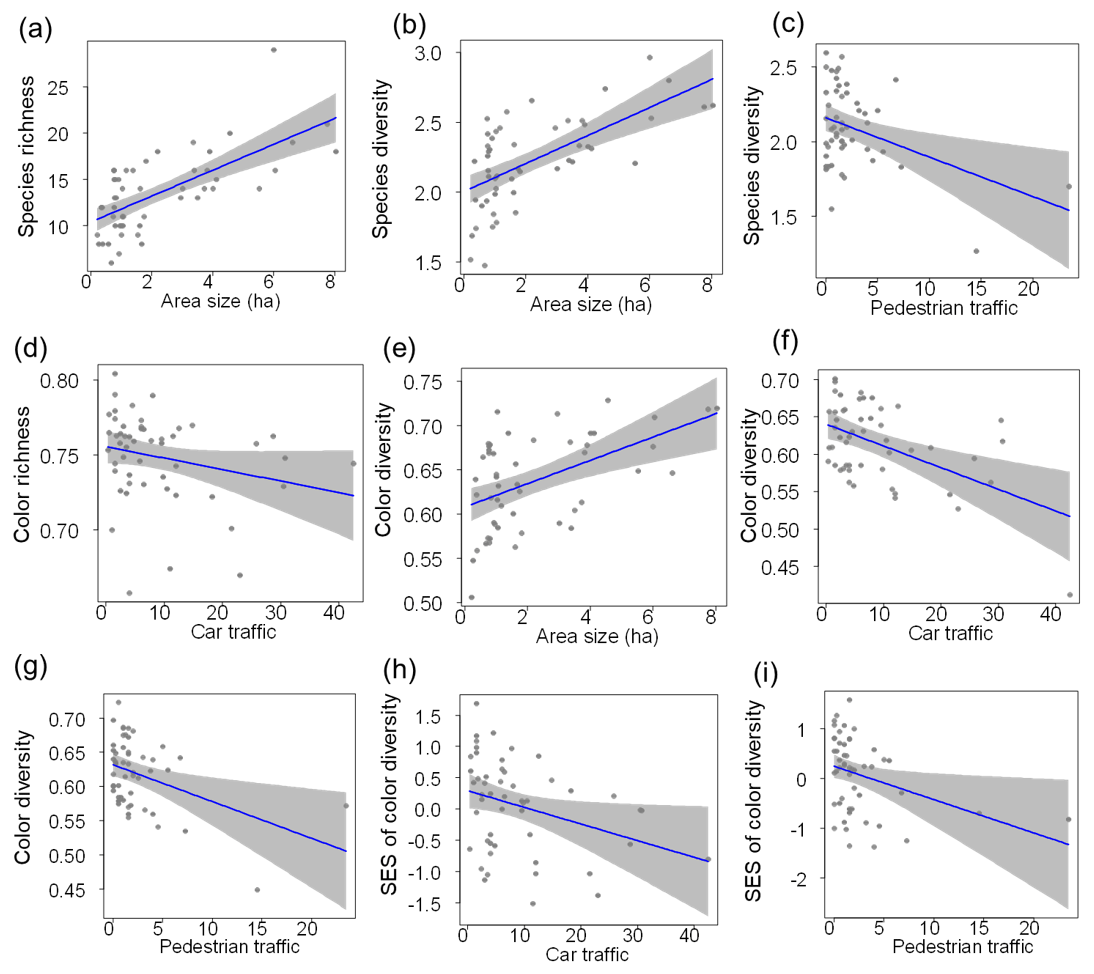
Figure 2Final generalized linear models showing the relationships between environmental variables and bird (a) taxonomic species richness, (b, c) taxonomic Shannon diversity, (d) color richness, (e–g) color diversity, and (h, i) standardized effects sizes (SESs) of color diversity in urban parks of central Argentina. Blue lines indicate the fitted model, and grey bands indicate 95 % confidence intervals.
Bird color richness tended to be negatively related to car traffic (LRT = 0.03, P=0.082, r2=0.06; Table 2c; Fig. 2d). After accounting for species richness, bird color richness was not related to any environmental variable (LRT = 2.18, P=0.147; Table 2d). Thus, bird color richness was random in urban parks. Bird color diversity was positively related to green-area size and negatively associated with car and pedestrian traffic (LRT = 0.11, P<0.001, r2=0.52; Table 2e; Fig. 2e–g). After accounting for species richness, bird color diversity was negatively related to car and pedestrian traffic only (LRT = 9.84, P<0.001, r2=0.45; Table 2f; Fig. 2h, i). Therefore, green areas with the highest car and pedestrian traffic had a clustered color composition.
Bird color composition changed with the environmental characteristics of green areas (Fig. 3). The number of grey and sexually dimorphic birds increased in green areas near the urban center. The number of brown, grey, and iridescent birds increased with increasing green-area size, whereas yellow and blue birds increased in green areas near the rural areas. Black birds tended to be negatively related to habitat diversity.
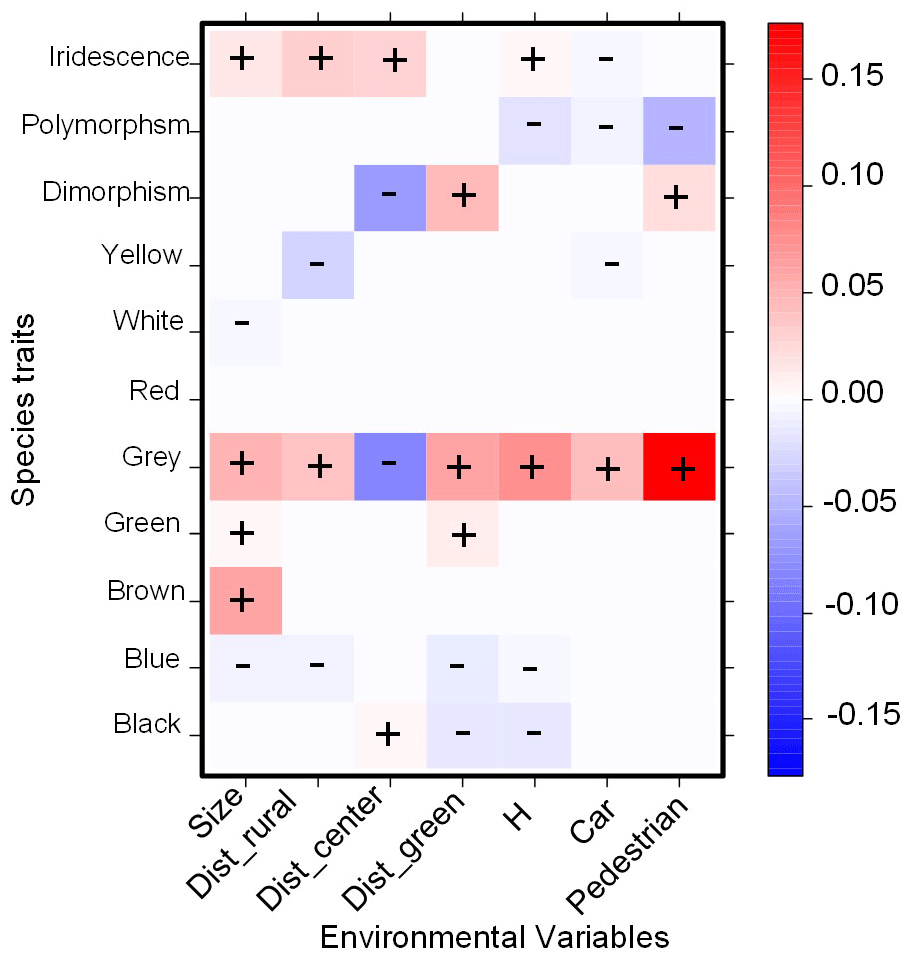
Figure 3Relationship between bird color composition and environmental variables of urban parks in six cities of central Argentina. Standardized coefficients for all environment–trait interaction terms are presented, which represent the interaction between the matrices of species × sites, environment × sites, and species traits × species (Brown et al., 2014). Coefficients were obtained with the least absolute shrinkage and selection operator (LASSO; Brown et al., 2014) with red colors indicating positive relationships and blue colors indicating negative relationships. Dist_rural: minimum distance to rural areas; Dist_center: minimum distance to urban center; Dist_green: minimum distance to other green areas; H: habitat diversity; Size: size of urban park; Car: number of cars passing on the street closest to the urban park; Pedestrian: number of people at each urban park.
As expected, bird taxonomic richness and diversity increased with area size and decreased with increasing human disturbance. However, color diversity was positively related to green-area size only when not corrected by species richness, and it showed a negative relationship with human disturbance. Therefore, the increase in green-area size is expected to be associated with an increase in bird taxonomic and color diversity, but human disturbance is expected to filter bird colors. Indeed, green areas near urban centers and with the highest human disturbance were dominated by grey and sexually dimorphic species.
Green-area size positively affected the bird taxonomic diversity. This result agrees with theoretical expectations (Arrhenius, 1921; MacArthur and Wilson, 1967). The highest bird diversity in large green areas could be related to decreased extinction risk (MacArthur and Wilson, 1967; Dri et al., 2021). Although green-area isolation was expected to affect species diversity (MacArthur and Wilson, 1967; Fernández-Juricic, 2000), our results showed no relationship between species diversity and distance to rural areas. Other studies conducted in urban green areas also showed that isolation was not an important factor in molding species diversity (Beninde et al., 2015; Leveau et al., 2019), suggesting that the impervious surface is not a significant dispersion barrier for bird species in the regional pool.
The patterns of color richness and diversity along urban parks varied whether they were corrected for species richness or not. Color richness tended to be negatively related to car traffic. However, richness-corrected values did not correlate with any environmental variable. Therefore, the loss of color richness due to car traffic seems to be dependent on the patterns of species richness. On the other hand, color diversity increased with green-area size, but the richness-corrected values did not correlate with area size. Thus, the relationship between green-area size and color diversity seems to be mediated by changes in species richness, and probably no process of species color filtering due to area size occurs.
The negative relationship between color diversity and car and pedestrian traffic was maintained when species richness was taken into account. Therefore, these disturbances probably acted as a filter of species colors in urban parks. On the one hand, car traffic could be acting indirectly on bird color diversity by excluding bird species due to noise (Francis et al., 2011), except those species adapted to noise that also are grey species. On the other hand, the effect of pedestrians could also act indirectly on bird color diversity. Pedestrian traffic and the food pedestrians could provide to birds may favor the abundance of species with few colors, for example, grey in the case of the house sparrow (Passer domesticus), the eared dove (Zenaida auriculata), and the picazuro pigeon (Patagioenas picazuro) (Leveau, 2019; Hambuckers et al., 2023; Swartz et al., 2023).
Bird color composition changed in relation to environmental variables of green areas. The abundance of species with grey plumage was positively correlated with pedestrian and car traffic, habitat diversity, area size, and distance to rural areas (city size) and increased near the urban center. Species with grey plumage that may become abundant in green areas are often omnivorous or granivorous species that form flocks to feed on food discarded by humans (Jokimäki and Suhonen, 1998; Leveau, 2013; Patankar et al., 2021). Such species, like the house sparrow and the eared dove, are adapted to disturbed sites (Baptista et al., 2020; Lowther and Cink, 2020) which have high levels of noise due to car traffic. In addition, the dominance of species with grey plumage was related to the distance to rural areas, which is an indicator of city size. City size is positively related to people density and the level of urbanization surrounding green areas, which may favor the abundance of grey species.
Yellow and blue birds tended to be more abundant near rural areas or in small cities. The closeness of rural areas may favor the dispersion of individuals and the availability of resources near urban parks. The relationship between city size and bird colors supports findings made by Leveau (2019) along impervious surface gradients in three of the six cities analyzed in this study (Balcarce, Mar del Plata, and Miramar).
Camouflage and the avoidance of predation have been postulated as the main factors shaping animal colors in anthropogenic landscapes (Delhey and Peters, 2017). It is expected that birds will choose surfaces that match their plumage color (Endler, 1993). In urban environments dominated by impervious grey surfaces, grey plumage may favor camouflage to avoid humans and other common mammals such as cats and dogs (Møller and Ibáñez-Álamo, 2012; Morelli et al., 2023). However, urban parks are usually dominated by green surfaces composed of lawns where grey birds feed. The mismatch between the green surface and the grey plumage can be offset by flocking, which is an antipredator strategy (Beauchamp, 2023).
The loss of bird color diversity in urban parks may be related to a loss of cultural services. Bird color has been described as one feature appreciated by humans (Lišková et al., 2015; Belaire et al., 2015). In addition, particular colors such as yellow and blue that were described as preferred by humans (Lišková et al., 2015; Correia et al., 2016; Garnett et al., 2018) declined with city size and distance to rural areas in our study. Therefore, local factors such as human disturbance and landscape factors such as closeness to rural areas should be controlled to avoid the loss of cultural ecosystem services in urban parks.
The results obtained showed that bird taxonomic diversity and color diversity were related to green-area size and human disturbance. After accounting for species richness, bird color diversity was mainly related to human disturbance. Therefore, the increase in color diversity with area seems to be mediated by an increase in species richness. Bird color composition changed according to green-area size, disturbance, park isolation, and city size. Grey species were more common near the urban center of cities, whereas yellow birds were more common in parks near rural areas. Bird color composition in urban parks seems to be influenced by biotic interactions between birds and humans, mediated by the availability of food resources, and the proximity to highly urbanized areas. Finally, the relations between color diversity and composition with green-area size and human disturbance suggest a loss of ecosystem services. Future studies should focus on temporal dynamics of color diversity in urban green areas, such as seasonal variations (Leveau, 2022), and the inclusion of other bird traits related to cultural ecosystem services, such as song diversity (Goodness et al., 2016; Han et al., 2022).
The dataset is available as the Supplement.
The supplement related to this article is available online at: https://doi.org/10.5194/we-24-1-2024-supplement.
Both authors have made substantial contributions in terms of conception, design, acquisition of data, analysis, and interpretation. Moreover, both authors were involved in the drafting and approval of the final version of the paper submitted.
The contact author has declared that neither of the authors has any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
We thank Sonja Knapp, Adrien Frantz, and the anonymous reviewer for their useful comments that helped to improve the quality of the manuscript.
This research has been supported by the Agencia Nacional de Promoción de la Investigación, el Desarrollo Tecnológico y la Innovación (grant no. PICT 2015-0978).
This paper was edited by Sonja Knapp and reviewed by Adrien Frantz and one anonymous referee.
Andrade, R., Larson, K. L., Franklin, J., Lerman, S. B., Bateman, H. L., and Warren, P. S.: Species traits explain public perceptions of human–bird interactions, Ecol. Appl., 32, e2676, https://doi.org/10.1002/eap.2676, 2022.
Arrhenius, O.: Species and area, J. Ecol., 9, 95–99, 1921.
Bailey, S. F.: Latitudinal gradients in colors and patterns of passerine birds, Condor, 80, 372–381, 1978.
Baptista, L. F., Trail, P. W., Horblit, H. M., Boesman, P. F. D., and Garcia, E. F. J.: Eared Dove (Zenaida auriculata), version 1.0, in: Birds of the World, edited by: del Hoyo, J., Elliott, A., Sargatal, J., Christie, D. A., and de Juana, E., Cornell Lab of Ornithology, Ithaca, NY, USA, https://doi.org/10.2173/bow.eardov1.01, 2020.
Beauchamp, G.: Susceptibility to Predation Varies with Body Mass, Foraging Niche, and Anti-Predator Responses among Bird Species, Birds, 4, 73–84, 2023.
Belaire, J. A., Westphal, L. M., Whelan, C. J., and Minor, E. S.: Urban residents' perceptions of birds in the neighborhood: Biodiversity, cultural ecosystem services, and disservices, Condor, 117, 192–202, 2015.
Beninde, J., Veith, M., and Hochkirch, A.: Biodiversity in cities needs space: a meta-analysis of factors determining intra-urban biodiversity variation, Ecol. Lett., 18, 581–592, 2015.
Brown, A. M., Warton, D. I., Andrew, N. R., Binns, M., Cassis, G., and Gibb, H.: The fourth-corner solution–using predictive models to understand how species traits interact with the environment, Methods Ecol. Evol., 5, 344–352, 2014.
Butcher, G. S. and Rohwer, S.: The evolution of conspicuous and distinctive coloration for communication in birds, in: Current ornithology, Springer US, Boston, MA, 51–108 pp., https://doi.org/10.1007/978-1-4757-9918-7, 1989.
Colding, J., Gren, Å., and Barthel, S.: The incremental demise of urban green spaces, Land, 9, 162, https://doi.org/10.3390/land9050162, 2020.
Cooney, C. R., He, Y., Varley, Z. K., Nouri, L. O., Moody, C. J., Jardine, M. D., Liker, A., Székely, T., and Thomas, G. H.: Latitudinal gradients in avian colourfulness, Nat. Ecol. Evol., 6, 622–629, 2022.
Correia, R. A., Jepson, P. R., Malhado, A. C., and Ladle, R. J.: Familiarity breeds content: Assessing bird species popularity with culturomics, PeerJ, 4, e1728, https://doi.org/10.7717/peerj.1728, 2016.
Curzel, F. E., Bellocq, M. I., and Leveau, L. M.: Local and landscape features of wooded streets influenced bird taxonomic and functional diversity, Urban For. Urban Green., 66, 127369, https://doi.org/10.1016/j.ufug.2021.127369, 2021.
De Bello, F., Carmona, C. P., Dias, A. T., Götzenberger, L., Moretti, M., and Berg, M. P.: Handbook of trait-based ecology: from theory to R tools, Cambridge University Press, Cambridge, 295 pp., https://doi.org/10.1017/9781108628426, 2021.
Delhey, K. and Peters, A.: Conservation implications of anthropogenic impacts on visual communication and camouflage, Cons. Biol., 31, 30–39, 2017.
Dri, G. F., Fontana, C. S., and de Sales Dambros, C.: Estimating the impacts of habitat loss induced by urbanization on bird local extinctions, Biol. Conserv., 256, 109064, https://doi.org/10.1016/j.biocon.2021.109064, 2021.
Endler, J. A.: Some general comments on the evolution and design of animal communication systems, Philos. Trans. R. Soc. Lond. B, 340, 215–225, 1993.
Fernández-Juricic, E.: Bird community composition patterns in urban parks of Madrid: the role of age, size and isolation, Ecol. Res., 15, 373–383, 2000.
Fernández-Juricic, E.: Can human disturbance promote nestedness? A case study with breeding birds in urban habitat fragments, Oecologia, 131, 269–278, 2002.
Francis, C. D., Ortega, C. P., and Cruz, A.: Noise pollution filters bird communities based on vocal frequency, PLoS one, 6, e27052, https://doi.org/10.1371/journal.pone.0027052, 2011.
Frynta, D., Lišková, S., Bültmann, S., and Burda, H.: Being attractive brings advantages: the case of parrot species in captivity, PloS one, 5, e12568, https://doi.org/10.1371/journal.pone.0012568, 2010.
Garnett, S. T., Ainsworth, G. B., and Zander, K. K.: Are we choosing the right flag-ships? The bird species and traits Australians and most attractive, PloS one, 13, e0199253, https://doi.org/10.1371/journal.pone.0199253, 2018.
González-Oreja, J. A., De La Fuente-Díaz, A. A., Hernández-Santín, L., Bonache-Regidor, C., and Buzo-Franco, D.: Can human disturbance promote nestedness? Songbirds and noise in urban parks as a case study, Landsc. Urban Plan., 104, 9–18, 2012.
Goodness, J., Andersson, E., Anderson, P. M., and Elmqvist, T.: Exploring the links between functional traits and cultural ecosystem services to enhance urban ecosystem management, Ecol. Indic., 70, 597–605, 2016.
Haila, Y.: A conceptual genealogy of fragmentation research: from island biogeography to landscape ecology, Ecol. Appl., 12, 321–334, 2002.
Hambuckers, A., Delcourt, J., Leborgne, B., and Cahill, J. R.: Artificial Green Corridors in an Andean City as Effective Support of Avian Diversity, Diversity, 15, 302, https://doi.org/10.3390/d15020302, 2023.
Han, P., Zhao, Y., Kang, Y., Ding, P., and Si, X.: Island biogeography of soundscapes: Island area shapes spatial patterns of avian acoustic diversity, J. Biogeogr., https://doi.org/10.1111/jbi.14551, online first, 2022.
Hartig, F.: Package “DHARMa”, R package, CRAN, https://cran.r-project.org/web/packages/DHARMa/index.html (last access: 18 March 2023), 2021.
Jokimäki, J. and Suhonen, J.: Distribution and habitat selection of wintering birds in urban environments, Landsc. Urban. Plan., 39, 253–263, 1998.
Kembel, S. W., Cowan, P. D., Helmus. M. R., Cornwell. W. K., Morlon. H., Ackerly. D. D., Blomberg. S. P., and Webb. C. O.: Picante: R tools for integrating phylogenies and ecology, Bioinformatics 26, 1463–1464, 2010.
Leveau, L. M.: Bird traits in urban–rural gradients: how many functional groups are there?, J. Ornithol., 154, 655–662, 2013.
Leveau, L. M.: Los efectos de la vegetación y temperatura sobre los ensambles de aves en gradientes urbano-rurales, Doctoral dissertation, PhD thesis, Universidad Nacional de Mar del Plata, Mar del Plata, 204 pp., 2014.
Leveau, L. M.: Urbanization induces bird color homogenization, Landsc. Urban. Plan., 192, 103645, https://doi.org/10.1016/j.landurbplan.2019.103645, 2019.
Leveau, L. M.: United colours of the city: A review about urbanisation impact on animal colours, Austral Ecol., 46, 670–679, 2021.
Leveau, L. M.: Does spring bring colors? Seasonal bird plumage color variation along urban-rural gradients, Perspect. Ecol. Conserv., 20, 103–110, 2022.
Leveau, L. M. and Ibáñez, I.: Nesting site and plumage color are the main traits associated with bird species presence in urban areas, Animals, 12, 1148, https://doi.org/10.3390/ani12091148, 2022.
Leveau, L. M. and Leveau, C. M.: Does urbanization affect the seasonal dynamics of bird communities in urban parks?, Urban Ecosyst., 19, 631–647, 2016.
Leveau, L. M., Ruggiero, A., Matthews, T. J., and Bellocq, M. I.: A global consistent positive effect of urban green area size on bird richness, Avian Res., 10, 1–14, 2019.
Linares, S., Iturrioz, I., and Di Nucci, J.: La expansión urbana de la ciudad de Tandil 1991–2020 mediante el uso del Landscape Expansion Index (LEI), Pleamar, Revista del Departamento de Geografía. Facultad de Humanidades, Universidad Nacional de Mar del Plata, 2, 59–77, 2022.
Lišková, S. and Frynta, D.: What determines bird beauty in human eyes?, Anthrozoös, 26, 27–41, 2013.
Lišková, S., Landová, E., and Frynta, D.: Human preferences for colorful birds: Vivid colors or pattern?, Evol. Psychol., 13, 147470491501300203, https://doi.org/10.1177/147470491501300203, 2015.
Liu, J., Bai, H., Ma, H., and Feng, G.: Bird diversity in Chinese urban parks was more associated with natural factors than anthropogenic factors, Urban Forest Urban Green, 43, 126358, https://doi.org/10.1016/j.ufug.2019.06.001, 2019.
Lopez, B. E., Urban, D., and White, P. S.: Testing the effects of four urbanization filters on forest plant taxonomic, functional, and phylogenetic diversity, Ecol. Appl., 28, 2197–2205, 2018.
Lowther, P. E. and Cink, C. L.: House Sparrow (Passer domesticus), version 1.0, in: Birds of the World, edited by: Billerman, S. M., Cornell Lab of Ornithology, Ithaca, NY, USA, https://doi.org/10.2173/bow.houspa.01, 2020.
MacArthur, R. H. and MacArthur, J. W.: On bird species diversity, Ecology, 42, 594–598, 1961.
MacArthur, R. H. and Wilson, E. O.: The theory of island biogeography. Princeton: Princeton University Press, 224 pp., ISBN 9780691088365, 1967.
Mao, Q., Sun, J., Deng, Y., Wu, Z., and Bai, H.: Assessing Effects of Multi-Scale Landscape Pattern and Habitats Attributes on Taxonomic and Functional Diversity of Urban River Birds, Diversity, 15, 486, https://doi.org/10.3390/d15040486, 2023.
Marzluff, J. M., Bowman, R., and Donnelly, R.: A historical perspective on urban bird research: trends, terms, and approaches, Avian ecology and conservation in an urbanizing world, Springer Science, New York, 585 pp., https://doi.org/10.1007/978-1-4615-1531-9, 2001.
Matthews, T. J., Guilhaumon, F., Triantis, K. A., Borregaard, M. K., and Whittaker, R. J.: On the form of species–area relationships in habitat islands and true islands, Glob. Ecol. Biogeogr., 25, 847–858, 2016.
Matthews, T. J., Wayman, J. P., Whittaker, R. J., Cardoso, P., Hume, J. P., Sayol, F., Proios, K., Martin, T. E., Baiser, B., Borges, P. A. V., Kubota, Y., dos Anjos, L., Tobias, J. A., Soares, F. C., Si, X., Ding, P., Mendenhall, C. D., Sin, Y. C. K., Rheindt, F. E., Triantis, K. A., Guilhaumon, F., Watson, D. M., Brotons, D. M., Battisti, D., Chu, O., and Rigal, F.: A global analysis of avian island diversity–area relationships in the Anthropocene, Ecol. Lett., 26, 965–982, https://doi.org/10.1111/ele.14203, 2023.
Melo, M. A., Campos-Silva, L. A., and Piratelli, A. J.: Red clay roof and NDVI drive changes in bird species composition and functional evenness in housing areas of São Paulo megacity, Brazil, Hornero, 37, 87–103, https://doi.org/10.56178/eh.v37i2.398, 2022.
Merlotto, A., Piccolo, M. C., and Bértola, G. R.: Crecimiento urbano y cambios del uso/cobertura del suelo en las ciudades de Necochea y Quequén, Buenos Aires, Argentina, Rev. de Geogr. Norte Gd., 53, 159–176, 2012.
Morelli, F., Benedetti, Y., Ibáñez-Álamo, J. D., Jokimäki, J., Mänd, R., Tryjanowski, P., and Møller, A. P.: Evidence of evolutionary homogenization of bird communities in urban environments across Europe, Glob. Ecol. Biogeogr., 25, 1284–1293, 2016
Morelli, F., Benedetti, Y., Su, T., Zhou, B., Moravec, D., Šímová, P., and Liang, W.: Taxonomic diversity, functional diversity and evolutionary uniqueness in bird communities of Beijing's urban parks: effects of land use and vegetation structure, Urban For. Urban Green., 23, 84–92, 2017.
Morelli, F., Leveau, L. M., Mikula, P., MacGregor-Fors, I., Bocelli, M. L., Quesada-Acuña, S. G., González-Lagos, C., Guitérrez-Tapia, P., Dri, G. F., Delgado-V., C., Garitano Zavala, A., Campos, J., Ortega Alvarez, R., Contreras Rodríguez, A. I., Souza López, D., Toledo, M. C. B., Sarquis, A., Giraudo, A., Echevarria, A. L., Fanjul, M. E., Martínez, M. V., Haedo, J., Cano Sanz, L. G., Peña Dominguez, Y. A., Fernandez, V., Marinero, V., Abilhoa, V., Amorin, R., Fonatana, C. S., da Silva, T. W., Zalewski Vargas, S. S., Escobar Ibáñez, J. F., Juri, M. D., Camín, S. R., Marone, L., Piratelli, A. J., Franchin, A. G., Crispim, L., Benitez, J., and Benedetti, Y.: Are birds more afraid in urban parks or cemeteries? A Latin American study contrasts with results from Europe, Sci. Total Environ., 861, 160534, https://doi.org/10.1016/j.scitotenv.2022.160534, 2023.
Møller, A. P. and Ibáñez-Álamo, J. D.: Escape behaviour of birds provides evidence of predation being involved in urbanization, Anim. Behav., 84, 341–348, 2012.
Nava-Diaz, R., Zuria, I., and Pineda-Lopez, R.: Taxonomic, phylogenetic and functional diversity of bird assemblages in urban green spaces: null model analyses, temporal variation and ecological drivers, Front. Ecol. Evol., 9, 795913, https://doi.org/10.3389/fevo.2021.795913, 2022.
Nielsen, A. B., Van Den Bosch, M., Maruthaveeran, S., and van den Bosch, C. K.: Species richness in urban parks and its drivers: A review of empirical evidence, Urban Ecosyst., 17, 305–327, 2014.
Oyarzabal, M., Clavijo, J., Oakley, L., Biganzoli, F., Tognetti, P., Barberis, I., Maturo, H. M., Aragon, R., Campanello, P. I., Prado, D., Oesterheld, M., and Leon, R. J. C.: Unidades de vegetación de la Argentina, Ecol. Austral, 28, 40–63, 2018.
Parodi, L. R.: La distribución geográfica de los talares en la provincia de Buenos Aires, Darwiniana, 4, 33–56, 1940.
Patankar, S., Jambhekar, R., Suryawanshi, K. R., and Nagendra, H.: Which traits influence bird survival in the city? A review, Land, 10, 92, https://doi.org/10.3390/land10020092, 2021.
Petchey, O. L., Evans, K. L., Fishburn, I. S., and Gaston, K. J.: Low functional diversity and no redundancy in British avian assemblages, J. Anim. Ecol., 76, 977–985, https://doi.org/10.1111/j.1365-2656.2007.01271.x, 2007.
R core Team: R: a Language and Environment for Statistical Computing (version 3.4.0), R Foundation for Statistical Computing, Vienna, Austria, https://www.r-project.org/ (last access: 25 September 2019), 2019.
Rangel, T. F., Diniz-Filho, J. A. F., and Bini, L. M.: SAM: a comprehensive application for spatial analysis in macroecology, Ecography, 33, 46–50, 2010.
Ricklefs, R. E. and Lovette, I. J.: The roles of island area per se and habitat diversity in the species–area relationships of four Lesser Antillean faunal groups, J. Anim. Ecol., 68, 1142–1160, 1999.
Shannon, C. E.: A mathematical theory of communication, Bell Syst. Tech. J., 27, 379–423 and 623–656, 1948.
Simberloff, D.: Experimental zoogeography of islands: effects of island size, Ecology, 57, 629–648, 1976.
Swartz, T. M., Gleditsch, J. M., and Behm, J. E.: A functional trait approach reveals the effects of landscape context on ecosystem services provided by urban birds, Landsc. Urban Plan., 234, 104724, https://doi.org/10.1016/j.landurbplan.2023.104724, 2023.
Tews, J., Brose, U., Grimm, V., Tielbörger, K., Wichmann, M. C., Schwager, M., and Jeltsch, F.: Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures, J. Biogeogr., 31, 79–92, 2004.
Turak, N., Monnier-Corbel, A., Gouret, M., and Frantz, A.: Urbanization shapes the relation between density and melanin-based colouration in bird communities, Oikos, 2022, e09313, https://doi.org/10.1111/oik.09313, 2022.
Valente-Neto, F., Roque, F. D. O., Pauliquevis, C. F., Oliveira, A. K. M. D., Provete, D. B., Szabo, J. K., and Souza, F. L.: Loss of cultural and functional diversity associated with birds across the urbanization gradient in a tropical city, Front. Ecol. Evol., 9, 615797, https://doi.org/10.3389/fevo.2021.615797, 2021.
Vatcheva, K. P., Lee, M., McCormick, J. B., and Rahbar, M. H.: Multicollinearity in regression analyses conducted in epidemiologic studies, Epidemiology 6, 1–20, https://doi.org/10.4172/2161-1165.1000227, 2016.
Wang, Y., Naumann, U., Eddelbuettel, D., Warton, D., Byrnes, J., Silva, S., Niku, J., Renner, I., and Wright, S.: Package “mvabund”, Statistical methods for analysing multivariate abundance data, CRAN, https://cran.r-project.org/web/packages/mvabund/index.html (last access: 18 March 2023), 2018.
Webb, C. O., Ackerly, D. D., McPeek, M. A., and Donoghue, M. J.: Phylogenies and community ecology, Annu. Rev. Ecol. Evol. Syst., 33, 475–505, 2002.
Yang, X., Tan, X., Chen, C., and Wang, Y.: The influence of urban park characteristics on bird diversity in Nanjing, China, Avian Res., 11, 1–9, 2020.
Zhang, D.: Package “rsq”, R-Squared and Related Measures, CRAN, https://cran.r-project.org/web/packages/rsq/rsq.pdf (last access: 8 September 2021), 2021.
Zhang, W., Liang, C., Liu, J., Si, X., and Feng, G.: Species richness, phylogenetic and functional structure of bird communities in Chinese university campuses are associated with divergent variables, Urban Ecosyst., 21, 1213–1225, 2018.
Zhou, D. and Chu, L. M.: How would size, age, human disturbance, and vegetation structure affect bird communities of urban parks in different seasons?, J. Ornithol., 153, 1101–1112, 2012.