the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 07 Oct 2025
| 07 Oct 2025
Urban wildlife monitoring using citizen science suggests that European hedgehogs and badgers select different habitats
Daniel Issel
Gregor Laaha
Johannes Laimighofer
Johann G. Zaller
Richard Zink
Daniel Dörler
Florian Heigl
Some wildlife species can successfully adapt to urban environments. To prevent potential conflict of these species with humans or their pets, a better understanding of the presence of urban wildlife is needed. However, traditional monitoring methods are often inadequate because many privately owned properties are inaccessible. In this study, we analyse reports of European hedgehogs (Erinaceus europaeus or E. roumanicus) and badgers (Meles meles) provided by two long-term citizen science projects in the city of Vienna, Austria – stadtwildtiere.at and roadkill.at – to assess habitat preferences and potential ecological interactions. Vienna has a human population of about 2×106 and covers an area of 415 km2, 50 % of which is green space in the form of forests, parks and private gardens. A total of 356 hedgehog and 918 badger sightings were reported between 2012 and 2023. Sightings of both species were positively associated with a mix of sealed/built-up areas and green spaces with meadows and shrubs. However, sightings of both species were negatively associated with arable land, most likely due to the avoidance of open terrain, reduced food availability or simply because both nocturnal species were more difficult to spot on dark arable land. The steeper the slope of a habitat, the fewer hedgehogs were reported, whereas for badgers, a positive correlation between slope and reports was observed in areas with built-up fractions over 15 %. Overall, we observed hardly any hedgehog reports in areas in which badgers were reported. We conclude that citizen science wildlife monitoring can be a good data source to better understand human–wildlife interactions and could therefore be a model for other urban areas and species.
- Article
(2324 KB) - Full-text XML
- BibTeX
- EndNote




Urbanization is an ongoing worldwide trend in human population distribution (United Nations, Department of Economic and Social Affairs, Population Division, 2019). However, urban areas have also become attractive wildlife habitats, especially for species exhibiting a high level of adaptability in the form of behavioural and dietary flexibility, as well as disturbance tolerance (Lowry et al., 2013; Santini et al., 2018). Examples in Europe are the red fox (Vulpes vulpes) (Walter et al., 2018), the European hedgehog (Erinaceus sp.) (Hubert et al., 2011; Williams et al., 2018b; van de Poel et al., 2015; Taucher et al., 2020) and the Eurasian badger (Meles meles) (Geiger et al., 2018; Huck et al., 2008; Lovell et al., 2022), which inhabit urban habitats given the wide availability of anthropogenic food sources (Harris, 1984; Hubert et al. 2011; Gazzard et al., 2022; Gomes et al., 2019), good conditions for hibernation and shelter (Hubert et al., 2011), lower abundance of predators (Williams et al., 2018a; Poel et al., 2015) and restricted hunting regulations. Some urban species (e.g. foxes and badgers) can cause conflicts with humans, their properties or their domestic animals (Araújo, 2003; Ives et al., 2016; Kettel et al., 2018).
Hedgehog densities are much higher in urban areas compared to rural areas (Hubert et al., 2011; Schaus et al., 2020). In urban areas, hedgehogs mainly use gardens (here defined as privately owned non-sealed areas around buildings) as their preferred habitat (App et al., 2022; Baker and Harris, 2007; Gazzard et al., 2022; Hof and Bright 2009; Williams et al., 2015; Williams et al., 2018a). Although there are no notable human–wildlife conflicts, hedgehog numbers have declined on a national scale in Great Britain (Matthews et al., 2018; Pettett et al., 2018) and in the city of Zurich, Switzerland (Taucher et al., 2020). Factors for the decline are habitat degradation (Taucher et al., 2020; Hof and Bright, 2009; Braaker et al., 2017; Pettett et al., 2018; Rondinini and Doncaster, 2002); road kills (Łopucki et al., 2021; Wright et al., 2020); lower food availability in the form of invertebrates, mainly through intense agriculture (Taucher et al., 2020; Rautio et al., 2015; Nottingham et al., 2019); and a high risk of predation, mostly by badgers (Poel et al., 2015; Williams et al., 2018a; Hof et al., 2012; Hof et al., 2019; Pettett et al., 2018). The International Union for Conservation of Nature (IUCN) therefore recently updated the status of E. europaeus to “Near Threatened” (Gazzard and Rasmussen, 2024).
The Eurasian badger is listed as “Least Concern” by the IUCN, due to increasing population sizes in most of its natural range (Kranz et al., 2016). The main reasons for increasing badger populations are the control of rabies, for which badgers can serve as a reservoir host (Schwierz and Wachendörfer, 1981; Smith and Wilkinson, 2002), and a diverse adaptable diet (Gomes et al., 2019; Van Den Berge et al., 2022). Badgers mainly use structure-rich habitats such as (the edges of) woodland (Feore and Montgomery, 1999; Huck et al., 2008; Piza-Roca et al., 2018) or shrubs (Davison et al., 2009; Harris, 1984; Huck et al., 2008) due to their high cover and structural support of roots for their setts (i.e. burrows) and high abundances of invertebrates as an important food source (Gomes et al., 2019; Kruuk and Parish, 1981; Van Den Berge et al., 2022). Moreover, badgers prefer slopes as their sett sites (Fischer and Weber 2003; Huck et al., 2008; Macdonald et al., 2004; Reid et al., 2012). Badger activity declines when humans are present (Lovell et al., 2022; Schley et al., 2004); however, an increase in their activity in the city of Zurich was observed (Geiger et al., 2018).
Hedgehogs and badgers have a complex ecological relationship. Both species show similar diets, feeding mainly on invertebrates (Gomes et al., 2019; Nottingham et al., 2019), and thus compete for food. In areas where both species are present, lower abundances of invertebrates are observed (Hof et al., 2012). Moreover, badgers are known as the main predator of hedgehogs, and hedgehogs are avoiding areas with high badger abundances (Hof, 2009; Hof et al., 2019; Pettett et al., 2018, van de Poel et al., 2015; Williams et al., 2018a; Young et al., 2006). Additionally, hedgehogs prefer more cover structures like field margins and hedgerows (Hof et al., 2012; Pettett et al., 2017) when badgers are present (Doncaster, 1992; Hof et al., 2012; Young et al., 2006; Pettett et al., 2017; Williams et al., 2018a; Poel et al., 2015). In extreme situations, the interspecific interactions described can lead to local extinctions of hedgehogs in areas with high numbers of badgers (Micol et al., 1994; Young et al., 2006).
Monitoring urban populations can be challenging, because the large number of privately owned properties with prohibited access impair the implementation of traditional research methods. Therefore, citizen science approaches became popular, with reported sightings of animal species containing important meta information such as date, time and exact location of the sighting. These datasets are often presence only, lacking information on areas where species are absent, and they frequently exhibit uneven spatial and temporal coverage. To address these challenges, various statistical methods have been employed, including conditional probabilities (Heigl et al., 2017; Walter et al., 2018) and generalized linear models (GLMs) (Tiago et al., 2017; Goldstein and de Valpine, 2022). Conditional probabilities are particularly effective for examining isolated associations, such as the relationship between species presence and specific land-use classes, due to their simplicity and minimal data preparation requirements. GLMs, in contrast, provide a more flexible framework for incorporating multiple predictors and exploring complex interactions. These complementary strengths make both methods valuable tools for addressing the challenges of analysing citizen science data.
Here, we analysed data from long-established citizen science projects for hedgehogs and badgers in the city of Vienna, Austria, to investigate the following research questions. (i) Are hedgehogs and badgers more likely to be reported in land-use classes typical for private gardens, such as herb and meadow areas and shrub layers? (ii) Does a sloped terrain favour reports of badgers due to their preference for sett locations (Huck et al., 2008)? (iii) Are conditional probabilities a valid method to analyse the effect of land-use classes on hedgehog and badger reports in Vienna? We hypothesize that both species are reported mostly in areas close to private gardens, given a combination of food availability (Gazzard et al., 2022; Gomes et al., 2019) and good detectability by participants. Moreover, we also hypothesize that badgers are more likely to be reported from areas with higher slopes than hedgehogs, due to their sett locations (Huck et al., 2008). Finally, we hypothesize that conditional probabilities show similar results compared to alternative statistical methods (negative binomial models) and, therefore, are a valid method for analysing wildlife reports (Heigl et al., 2017; Walter et al., 2018).
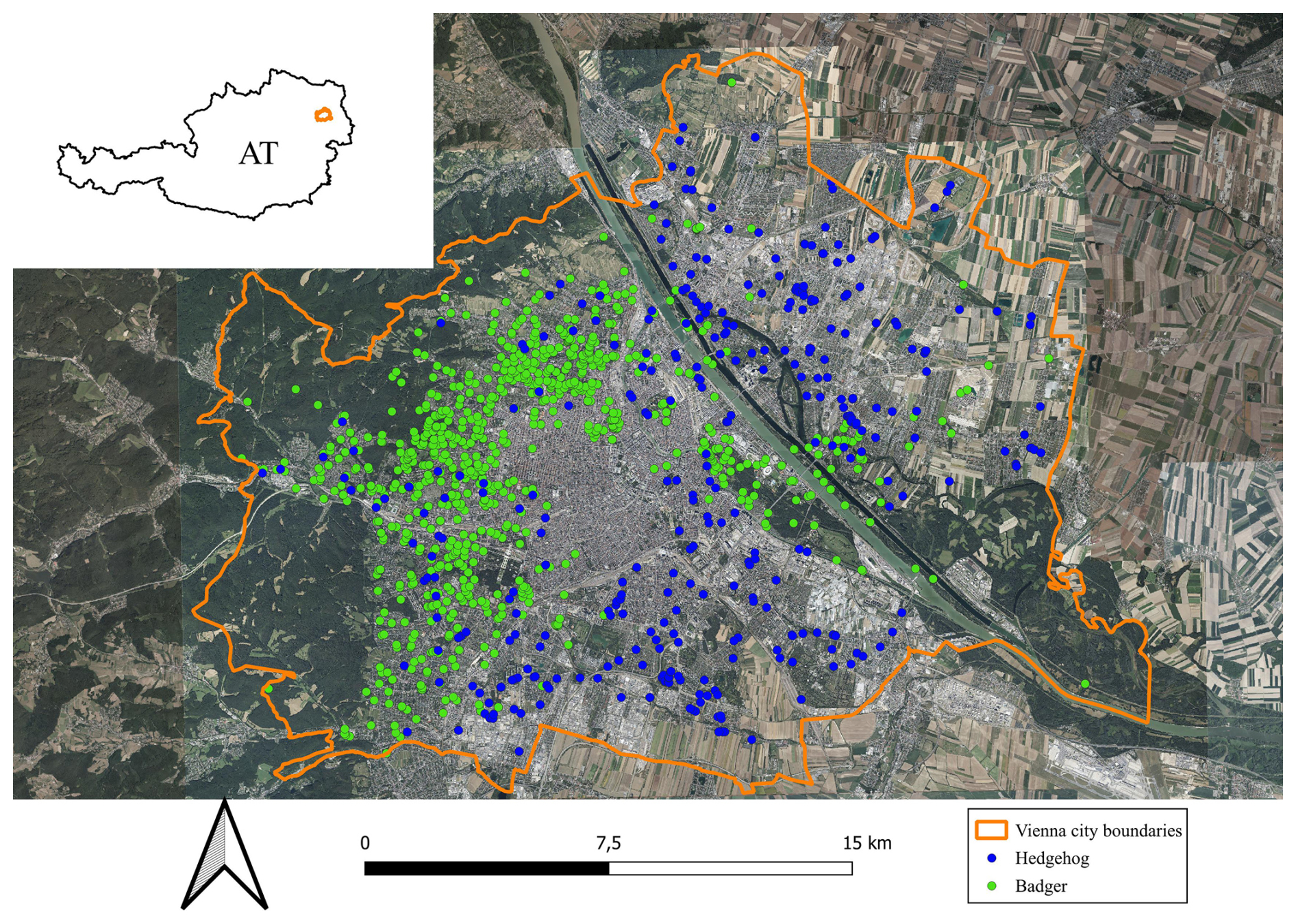
Figure 1Satellite image of the study area, the city of Vienna. Blue (hedgehog) and green (badger) points show reported locations between 2012 and 2023. Source: © https://basemap.at (last access: 26 February 2025)
2.1 Study area
We conducted the study in the city of Vienna, Austria (city centre coordinates: 48°12′ N, 16°22′ E), determined by its administrative borders. Vienna is the capital of Austria with about 2×106 inhabitants, an area of 414 km2 and an elevation gradient of 151–544 m above sea level. The city has a temperate climate with an annual mean temperature of 10.9 °C and an average annual rainfall of 703 mm. Over 50 % of the city area is classified as “green space”, containing forests, parks, arable land and gardens (Haberfellner et al., 2022). However, the green space is unevenly distributed, with highly sealed districts (up to 86 %) in the city centre and large green space areas towards the outer city borders (such as the Vienna Woods in the west, the national park Lobau in the east and arable land in the north and south; Fig. 1).
2.2 Hedgehog and badger reports
Two citizen science projects were used to obtain reports on hedgehog and badger sightings in Vienna. Participants reported road-killed vertebrate species to the project Roadkill (https://roadkill.at/, last access: 26 February 2025) and sightings of all wildlife, dead or alive, within the city of Vienna to the project StadtWildTiere Wien (https://wien.stadtwildtiere.at/, last access: 26 February 2025). Both projects enable participants to report their sightings – including date, time and coordinates – via a website or a mobile app. StadtWildTiere and Roadkill have a similar quality assurance system (Heigl et al., 2022; Zink and Walter, 2016). For this study, we only used the highest possible validation rank where experts validate reports based on provided photos and overall plausibility. If several reports of the same species came from the same location (equal GPS coordinates) within 72 h, we excluded all reports after the first one to avoid multiple reports of the same individual. Therefore, we used 356 hedgehog reports, 74 from project Roadkill and 282 from StadtWildTiere; and 918 badger reports, two from Roadkill and 916 from StadtWildTiere. Roadkill provided data from 2014 to 2023 and StadtWildTiere from 2012 to 2023. Participants reported hedgehogs from 26 May 2012 to 3 July 2023 and badgers from 14 February 2015 to 8 June 2023.
2.3 Land-use data
The Municipal Department 22 – Environmental Protection in Vienna provides land-use classes for the entire area of Vienna. The data source is the Grünraummonitoring Wien 2018 (green space monitoring Vienna 2018) and contains raster data with a pixel size of 50 cm × 50 cm. The data consist of eight categories. Infrared aerial images were used to classify the first three categories by detecting vegetation in combination with relative height, which is the difference between a surface model and a terrain model.
-
Herb and meadow areas: height was < 1 m;
-
Shrub layers: height was between 1.1 and 4 m;
-
Treetop area: height was > 4 m.
A combination of infrared aerial images (again to detect vegetation) and the multi-purpose map of Vienna was used to classify the other categories.
-
Unsealed areas, e.g. water bodies, railroad lines;
-
Sealed areas, e.g. streets, parking lots or squares;
-
Built-up areas, e.g. buildings;
-
Green on built-up areas, e.g. roofs with vegetation;
-
Arable land, mostly areas designated for agricultural use, specifically for growing crops.
Other available land-use datasets are the zoning plan of Vienna (https://www.data.gv.at/katalog/de/dataset/flachenwidmungs-und-bebauungsplan-plandokumente-wien, last access: 26 February 2025) or OpenStreetMap (https://www.openstreetmap.org/, last access: 26 February 2025). However, the zoning plan often merges privately owned gardens with building areas, while OpenStreetMap categorizes gardens as green spaces, which also include parks and other meadow areas. Compared to these datasets, the Grünraummonitoring has a finer breakdown of green space into meadow areas, trees and shrub layers, making it the best available option for this study.
Additionally, we calculated the degree of slope inclination with data from the terrain model of the Municipal Department 41 – City Survey (https://www.data.gv.at/katalog/de/dataset/stadt-wien_gelndemodellrasterwien, last access: 26 February 2025) and the Quantum geographic information system (QGIS) algorithm slope (QGIS 3.32.3; QGIS Development Team, 2023). The algorithm calculated the change in elevation between each cell and its neighbouring cells of the terrain model.
2.4 Statistical analysis
We conducted all analyses using the statistical software R (version 4.3.0; R Core Team, 2023) in the integrated development environment RStudio (version 2024.4.2.764; RStudio Team, 2024) and Microsoft Excel for Microsoft 365 MSO (version 2501 Build 16.0.18429.20132) 64 bit (Microsoft Corporation, 2024).
We used Jaccard coefficients to calculate the geographic similarity between reports of both species by measuring the proportion of shared vs. unique elements (Eq. 1) (Heikinheimo et al., 2020; Levandowsky and Winter, 1971). We used a structure with squared grid cells of 16 ha, which we placed randomly over Vienna. We chose the cell size because it roughly matches the daily home ranges in urban environments for both study species (Korslund et al., 2023; Marco-Tresserras and López-Iborra 2022; Morris 1988; Šálek et al., 2015). For comparison, we calculated Jaccard coefficients (again via the grid structure) with the same number of randomly generated pseudo reports.
Equation (1): here we used the Jaccard coefficient between badger grids (B) and hedgehog grids (H), with B as the number of grid cells with a badger report and H as the number of grid cells with a hedgehog report.
Biologically, higher coefficients indicate greater overlap in the reported distributions of both species, while lower values suggest segregation. For Jaccard coefficients, we only used reports for 2016, because an equal report period allows for better interpretation of interspecific coexistence. In this year, both species had the highest number of reports. For all following calculations, we used the full dataset from 2012 to 2023.
To analyse the effect of land-use classes on reports of hedgehogs and badgers, we calculated conditional probabilities as described in Eq. (2) (Heigl et al., 2017; Walter et al., 2018). In the first step, we uploaded species observation points, land-use class data (European Petroleum Survey Group (EPSG): 32633) and the previously used 16 ha grid structure into QGIS.
Equation (2): this is the conditional probability of a species report (E) given a land-use class (D). ∑iAj is the sum of all fragments of a land-use class in present grids, and Atot is the total study area.
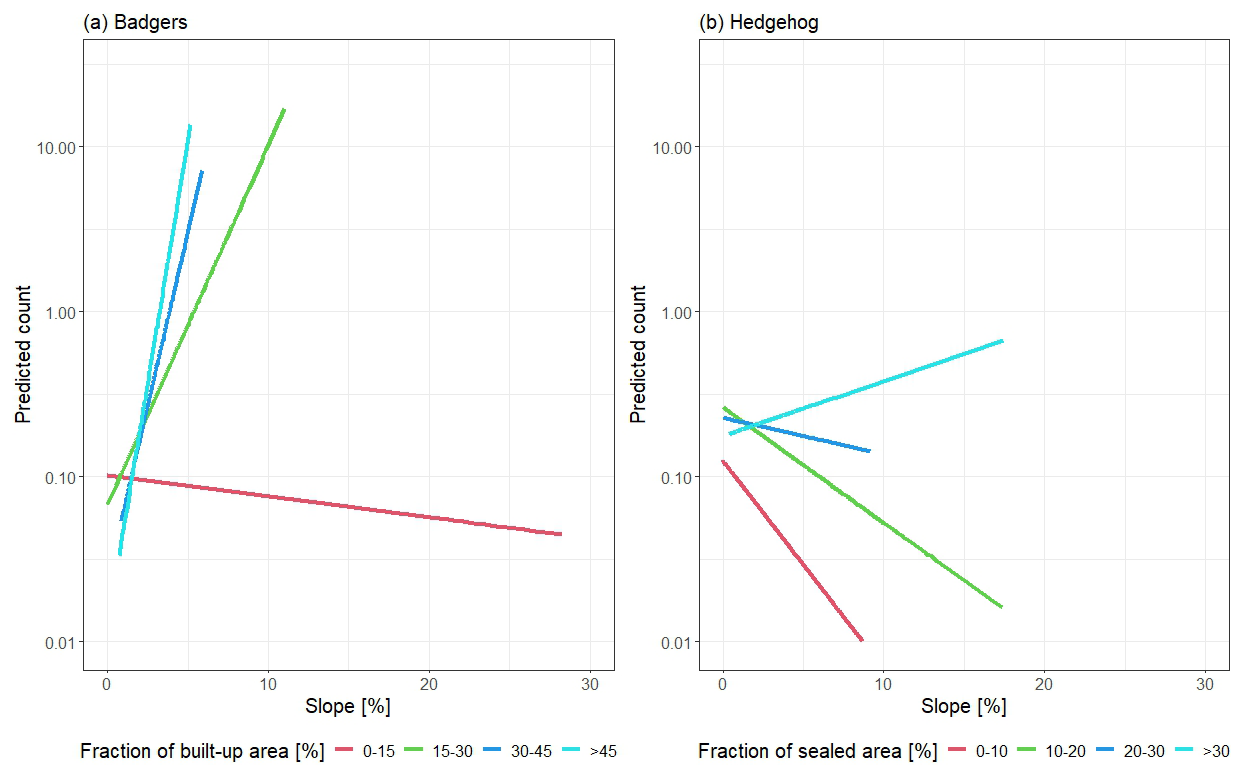
Figure 2Interaction plot between slope and fraction of (a) built-up areas for badgers and (b) sealed areas for hedgehogs. The y axis shows predicted counts.
In the next step, we extracted separately all grids with either a badger or hedgehog report and summarized all fragments of a land-use class in these present grids (). This sum was divided by the total study area (= size of Vienna, Atot). We then divided the total land-use class area by the total study area to get the overall probability of a land-use class P(Dj), which we then used as a divisor for the previously calculated quotient (P(E∩Dj). We used the algorithm raster layer unique values report to calculate area sizes. Lastly, we calculated P(E), which is the overall probability of a hedgehog or badger report, by dividing the area of presence grids by the total study area. When the conditional probability of a land-use class P(E|Dj) is lower than P(E), the land-use class hampers a report. If P(E|Dj) is higher than P(E), it favours a report.
As comparison to the conditional probabilities, we used GLMs to analyse the influence of land-use classes and slope inclination on reports. We used the number of reports in grid cells as the dependent variable and land-use classes and slope as independent variables. We coded land-use classes between 0 and 1, representing the area fraction of a land-use class in a grid cell and slope from 0 to 90, according to the mean slope degree of a grid cell. Because of the present distribution of the dependent variable with many zero counts (no badger/hedgehog report in that area), we used negative binomial models in the package MASS (Venables and Ripley, 2002). The models were tested for multi-collinearity by the variance inflation factor (VIF) with the package car (Fox and Weisberg, 2019), whereas the land-use classes Unsealed area and Green on built-up areas were removed due to VIF values being higher than 10 (Kutner et al. 2004). We also analysed models, examining the relationship between slope and land-use classes, as interactions may occur, such as fewer buildings being present in steep areas. Final models were selected through the use of stepwise variable selection (forward and backward) and the Akaike information criterion (AIC) as a metric. We generated interaction plots (Fig. 2) by dividing interacted land-use classes into four fractions depending on their percentage of area in the report grids (0 %–10 %, > 10 %, > 20 %, > 30 % for sealed areas (hedgehog); 0 %–15 %, > 15 %, > 30 %, > 45 % for built-up areas (badger)). We calculated the mean values for all covariates, except slope, for each land-use fraction. Then we predicted reports based on the fraction of the land-use class and varying slope values, while all other covariates were held constant. We used the packages ggplot2 (Wickham, 2016), tidyverse (Wickham et al., 2019) and gridExtra (Auguie, 2017).
3.1 Spatial distribution of hedgehog and badger reports
Hedgehogs and badgers were reported from different areas in Vienna. While hedgehogs were reported dominantly in the central and eastern parts of the city, badgers were more often reported in the western part of the city, near the Vienna Woods (Fig. 1).
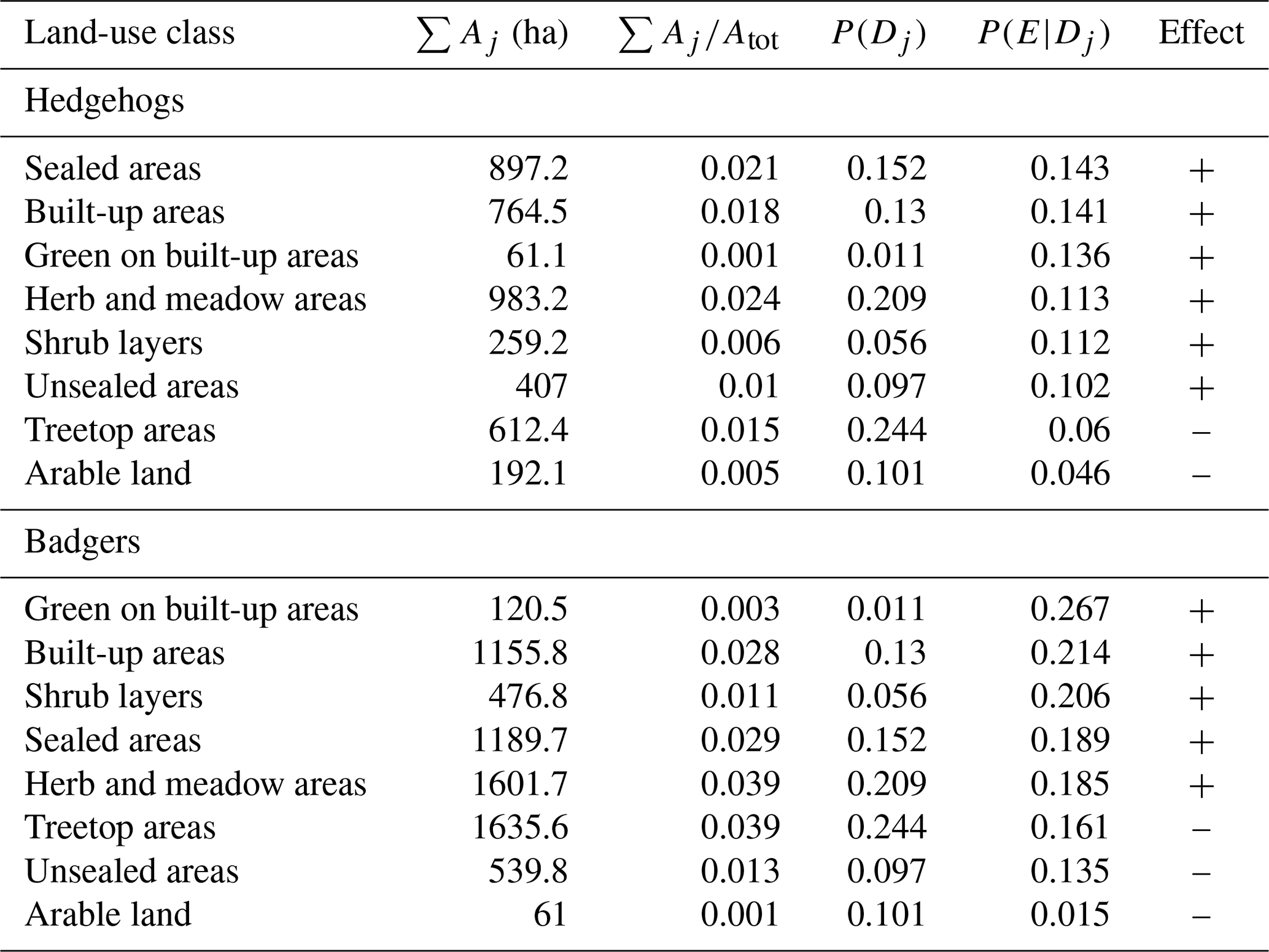
Table 1Land-use classes, their sizes in all grids with reports (∑ Aj), the factor when dividing ∑ Aj by the total area under investigation (∑ ), the overall probability of a land-use class P(Dj), the conditional probabilities P(E|Dj) and the effects of land-use classes on hedgehog and badger report probabilities. The threshold P(E) for a positive or negative effect is 0.1 for hedgehogs and 0.163 for badgers. Land-use classes are sorted by their P(E|Dj) value.
In 2016, hedgehogs were reported in 84 grids and badgers in 161 grids. The same number of randomly distributed pseudo points generated 107 grids for pseudo hedgehogs and 216 grids for pseudo badgers. With this information given, the Jaccard coefficient for hedgehog and badger grids in 2016 is = 0.008. In contrast, the Jaccard coefficient for the pseudo grids is = 0.042, which is 5.25 times higher than the Jaccard coefficient of the non-random grids. Therefore, the observed spatial distributions of hedgehog and badger reports are less overlapping than expected under random distribution.
3.2 Effect between land use and reports of hedgehogs and badgers
Effects of land use were analysed by calculating conditional probabilities for all eight land-use classes. The per cent distribution of the land-use classes (P(Dj)) over the city of Vienna was 24.4 % for treetop areas, 20.9 % for herb and meadow areas, 15.2 % for sealed areas, 13 % for built-up areas, 10.1 % for arable land, 9.7 % for unsealed areas, 5.6 % for shrub layers and 1 % for green on built-up areas.
The overall probability of grid cells with a hedgehog report P(E) was 10 %. The following land-use classes had a higher conditional probability P(E|Dj) than P(E) and therefore were positively associated with a hedgehog report: sealed areas, built-up areas, green on built-up areas, herb and meadow areas, shrub layers and unsealed areas. On the other hand, treetop areas and arable land were negatively associated with hedgehog reports. P(E) for badgers was 16.3 %. Green on built-up areas, built-up areas, shrub layers, sealed areas and herb and meadow areas were positively associated with badger reports. Treetop areas, unsealed areas and arable land hampered a badger report (Table 1).
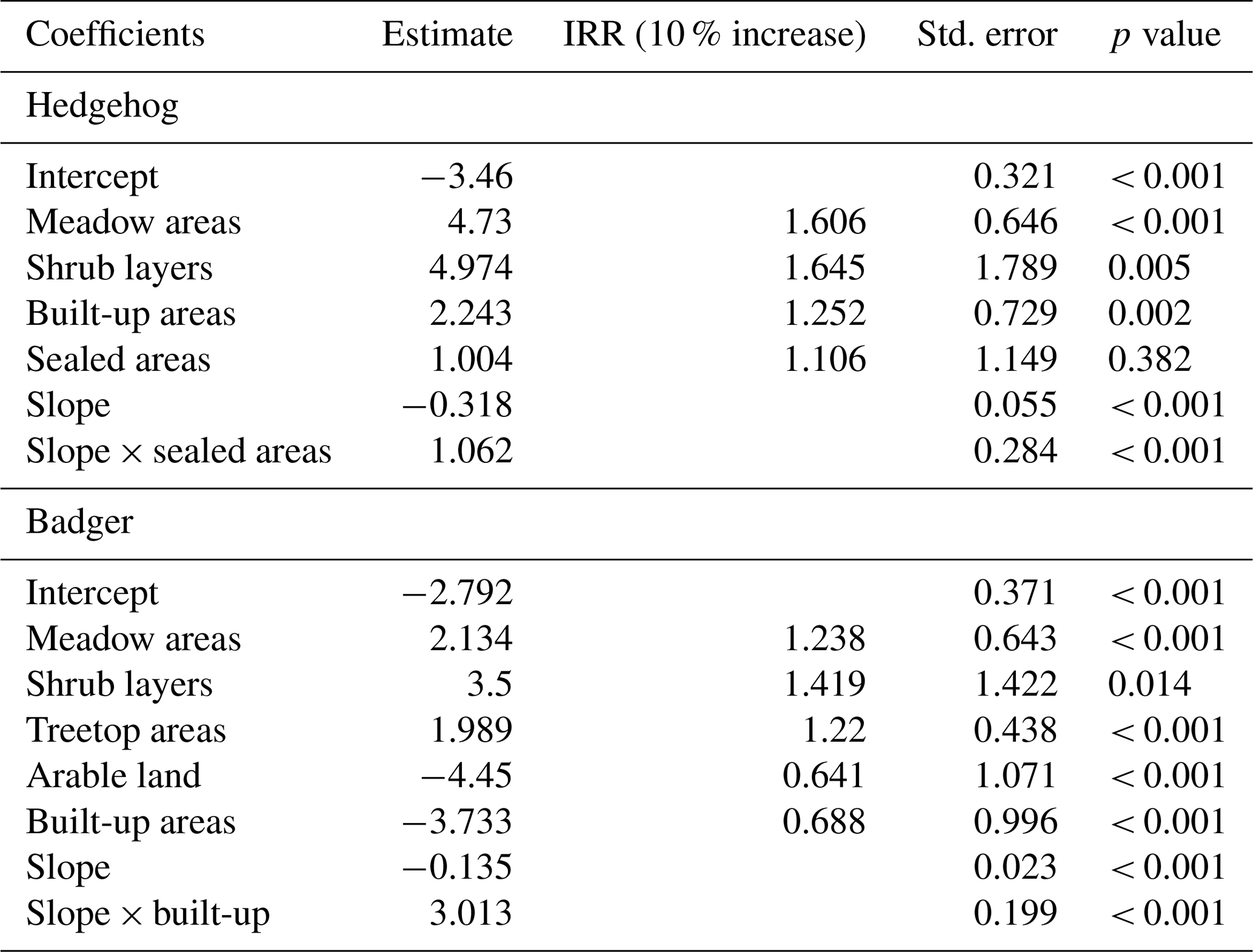
Table 2Results of the final negative binomial model for hedgehog and badger reports as dependent and land-use classes and slope as independent variables. The incidence rate ratio (IRR) shows the ecological effect sizes for land-use classes.
3.3 Model analysis of land-use class and slope
Non-interaction models had AIC values of 1980.3 in a model with all variables and 1976.8 after variable selection. These values were higher compared to the variable selected interaction model with an AIC of 1967.4. Therefore, the final hedgehog model included the variables meadow areas, shrub layers, built-up areas, sealed areas and slope and the interaction term of slope and sealed areas. Sealed areas was the only non-significant variable. While all other variables showed positive effects on reports of hedgehogs, slope had a negative effect (Table 2). The detailed analysis of the interaction plot in Fig. 2b showed a slight negative effect of slope and sealed fractions of 0 %–30 % and a small positive effect for fractions > 30 %. The range between different fraction categories are overall low with predicted hedgehog reports of less than one. All VIF values are < 10 (shrub layers: 1.451, meadow areas: 1.737, built-up areas: 2.11, sealed areas: 5.051, slope: 2.47, slope-sealed: 4.679), and the Cox–Snell pseudo R2 is 0.269.
The best-fitting non-interaction model had a higher AIC (3403.9) compared to the best-fitting interaction model with an AIC of 3208.5. Therefore, the final model for badgers contained the land-use classes of meadow areas, shrub layers, treetop areas, arable land, built-up areas and slope and the interaction term of slope and built-up areas. Variables of meadow areas, shrub layers and treetop areas and the interaction term showed positive effects on badger reports. Arable land, built-up areas and slope had negative effects. Figure 2a shows a high positive interaction between slope and fractions of built-up areas > 15 %, while fractions < 15 % had nearly no interaction effect on predicted badger reports. All VIF values are < 10 (meadow areas: 2.084, shrub layers: 1.393, treetop areas: 3.105, arable land: 1.142, built-up areas: 5.977, slope: 2.483, slope built-up: 3.605), and the Cox–Snell pseudo R2 is 0.348.
For a better ecological understanding, we included IRRs which show the change in report numbers on a 10 % increase of a land-use class (e.g. meadow areas = 1.238 = 23.81 % more reports; arable land = 0.641 = 35.9 % less reports).
The results of this investigation show that hedgehogs and badgers were reported from different parts of Vienna with different dominant land-use categories and slopes of habitats. The absence of mutual reports of hedgehogs and badgers could be one indicator of the predator avoidance theory. Previous studies found similar results, identifying badgers as the main predator of hedgehogs and showing that hedgehogs avoid areas with high badger densities (Hof, 2009; Hof et al., 2019; Pettett et al., 2018; van de Poel et al., 2015; Williams et al., 2018a; Young et al., 2006). However, we cannot conclude from the data that hedgehogs avoid badgers and were therefore reported in other areas. There could be other reasons for this, such as habitat suitability or the reporting behaviour of participants. Further investigations with additional parameters would be necessary to draw these conclusions.
Slope has a slight negative impact on hedgehog reports when sealed areas are absent or sparsely present. However, our data do not allow us to determine whether hedgehogs were reported in areas with low slopes because of the slope itself (Wright et al., 2020) or for other reasons. The positive interaction term between slope and sealed areas suggests that in steeper regions, the presence of sealed areas increases the likelihood of hedgehog reports, potentially due to higher human activity. Moreover, hedgehogs could prefer sealed areas in hilly regions to overcome steep slopes, as sealed areas are flattened. Nevertheless, as shown in Fig. 2b, the interaction term has overall low effects on reports, as the extreme scenarios (0 %–10 % sealed areas and > 25 % slope vs. > 30 % sealed areas and 1 % slope) have a difference of less than one report. Small effects of slope on badger reports contradict existing literature as slope is an important factor in badger sett location (Fischer and Weber 2003; Huck et al., 2008; Macdonald et al., 2004; Reid et al., 2012). However, this is only the case for areas with no or low fractions of built-up areas (0 %–15 %). We hypothesize that the low effect may be a reporting bias rather than ecological. Many participants are either not frequently present in steep areas or, if they are, it is primarily for activities like hiking, as the most hilly regions are located in the Vienna Woods. Since badgers are nocturnal, the likelihood of encountering them during daytime hiking is low, which could further amplify the reporting bias. Additionally, when hiking, badger encounters might be more expected compared to, for example, sightings in private gardens, potentially reducing the motivation to submit a report (Walter et al., 2018). This hypothesis is further supported by the positive interaction of areas with built-up fractions > 15 % that have high predicted counts (> 10) even on low slope values (< 10 %) (Fig. 2). Therefore, the interaction of increased human presence in hilly areas (through buildings), also during the night, combined with an ecological preference of badgers towards slopes, could make reports more likely.
In previous studies, both species showed a preference for private gardens (App et al., 2022; Davison et al., 2009; Dowding et al., 2010; Gazzard et al., 2022; Harris, 1984; Williams et al., 2018a). Hedgehogs often inhabit urban (private) gardens because of house owners providing pet food (Gazzard and Baker 2020; Gazzard et al., 2022; Hof and Bright, 2009; Hubert et al., 2011) and compost heaps (Gazzard et al., 2022; Taucher et al., 2020). Green space monitoring showed that two land-use classes dominated private gardens: meadow areas with herbaceous vegetation and shrub layers. Both land-use classes favoured a hedgehog report in the conditional probabilities and the GLM. Badgers showed similar results with positive effects on the likelihood of a report when herbaceous and meadow areas and shrub layers were dominant. This could be explained as badgers forage in private gardens, increasing their likelihood of being reported by participants (Davison et al., 2009; Harris, 1984), while also requiring structure-rich habitats (such as shrubs) for cover and structural support for their setts (Gomes et al., 2019; Huck et al., 2008; Van Den Berge et al., 2022). However, it remains unknown whether these two land-use classes have a positive effect on reports from private gardens, as herbaceous and meadow areas and shrub layers can also be found in other green spaces, e.g. parks.
Treetop areas (respectively the areas beneath) were negatively associated with hedgehog reports in the conditional probabilities and were excluded by the variable selection in the final hedgehog model. However, this result could be misinterpreted due to the underlying data structure, which did not distinguish between treetop areas in forests and treetop areas in other environments such as private gardens. There were no reports of hedgehogs in tree-rich areas of the Vienna Woods, the Lobau National Park or the Prater park area. Ecological reasons for this could be predator avoidance or lower habitat preference (App et al., 2022; Gazzard et al., 2022; Hof, 2009; Williams et al., 2018a; Young et al., 2006). Walter et al. (2018) had a similar trend for red foxes in Vienna and argued that the citizen science method could discriminate such areas, with animals present but not being reported because they are harder to detect, and participants would rather report wildlife in unexpected situations, such as in their private garden or in areas near their home, than in the forest or park, where they expect such animals. Therefore, large forest areas could lead to the misinterpretation of individual trees in gardens or smaller forest patches in urban environments (because they are in the same land-use class) which otherwise could favour a report, e.g. by providing an additional food source (Rautio et al., 2015). For badgers, (the edges of) woodland serves as a key urban habitat (Balestrieri et al., 2009; Balestrieri et al., 2016; Lovell et al., 2022), which may explain the positive effect on reports observed in the model. However, the underlying cause of the opposite negative effect in the conditional probabilities remains unclear. Additional research with a different, more precise selection of tree cover classes could be helpful in analysing the importance of urban trees for both species.
Arable land was negatively associated with a report for both species and was excluded by the AIC in the hedgehog model, which may have ecological and methodological reasons. It is known from previous studies that hedgehogs prefer lawns to arable land (Hubert et al., 2011) and, in urban habitats, private gardens (App et al., 2022; Williams et al., 2018a). Additionally, hedgehogs may avoid arable land due to high levels of agricultural intensification and low food availability caused by pesticides (Williams et al., 2018a). Badgers generally avoid open habitats such as arable land (Feore and Montgomery, 1999). Furthermore, reports from arable land appear to be methodologically improbable, given that participants are rarely present on such land, particularly in the evening and during the night, when in general most reports occurred.
Green on built-up areas had the lowest percentage of land cover of all land-use classes in Vienna (1 %). Moreover, the analysis of the data revealed that most of this land-use class consisted of trees overhanging neighbouring roofs and were therefore misclassified, probably due to an incorrectly programmed algorithm. Additionally, most areas of this specific land-use class are inaccessible for small- to medium-sized mammals due to their height. Therefore, the results of this land-use class have a low (ecological) explanatory value.
Sealed areas had positive effects on reports for both species in the conditional probabilities, but again, both GLMs showed no or only low effects. On the one hand, the presence of humans, who mainly use roads and paths in urban areas, parks or forests, can increase encounters and reports from these areas (Walter et al., 2018; Wine et al., 2015). Moreover, both species are known to be cultural successors that benefit from anthropogenic food sources (Gazzard and Baker, 2020; Gomes et al., 2019; Rautio et al., 2015), such as garbage cans, and may therefore prefer sealed areas up to a certain extent. On the other hand, traffic, especially in urban areas with high vehicle numbers, may reduce reports on sealed areas due to avoidance by hedgehogs and badgers (Dowding et al., 2010; Obidziński et al., 2013). Moreover, badger occurrence negatively correlates with human density (Schley et al., 2004), and badger activity decreases in the presence of humans (Lovell et al., 2022). These contrasting interpretations could account for the differing and non-significant analysis results. We balanced a possible bias towards sealed areas, also from the Roadkill dataset, by using the 16 ha grids.
Built-up areas were positively associated with reports of both species in the conditional probabilities and for hedgehogs in the GLM. This could once again be attributed to hedgehogs' preference for private human spaces, such as houses and gardens, where they benefit from resources like pet food provided by humans (Gazzard and Baker 2020; Gazzard et al., 2022; Hof and Bright, 2009; Hubert et al., 2011) and simply the higher presence of participants. Badgers showed, contrary to the conditional probabilities, a negative effect in the GLM for built-up areas, with no or low slope values (Fig. 2). Especially in the inner circle of Vienna, such landscape values are given, with nearly no badger reports (Fig. 1). These results are in contrast to a study from the city of Zurich that showed an increase of badgers in the whole city, including core areas (Geiger et al., 2018). Possible reasons for that may be again the avoidance of and reduced activity around humans (Lovell et al., 2022; Schley et al., 2004). Nevertheless, we have not used human population density as a variable because this information is only given on a district level, which is too imprecise for this analysis. Walter et al., (2018) used this factor for red fox reports in Vienna but found no significant effect. However, we expect a correlation between human density and the fraction of built-up areas.
Both methods of analysing land-use classes (conditional probabilities and GLMs) yielded similar outcomes regarding garden land classes, an important urban habitat for both species. In contrast, the contradictory results of treetop-, sealed- and built-up areas highlight the limitations of conditional probabilities in providing comprehensive insights. While conditional probabilities are particularly useful for a straightforward analysis of isolated associations like land-use data, they are less suited for study designs like ours that (have the data to) involve multiple variables. In such cases, GLMs are effective as they account for combined effects and provide uncertainty estimates, offering a more detailed understanding of the relationships between variables and species presence.
In conclusion, this study indicates that a mix of built-up and green space land-use classes favour citizen science reports of hedgehogs and badgers. This is most likely due to a mix of the ecological needs of the species and the employed citizen science method, which relies on data collectors who tend to be in urban areas close to built-up areas. Moreover, the results of this study revealed low effects of slope on reports of hedgehogs but positive effects on badger reports – if built-up areas are frequently present. Lastly, both species were mostly reported in different parts of Vienna, but it remains unclear whether this is due to predator avoidance or other factors. These results are predominantly in line with the current state of urban hedgehog and badger research, proving that citizen science can be an alternative to traditional research methods. However, this study also highlighted the shortcomings of citizen science, such as the reporting bias of humans, who are more likely to detect and report wildlife in unexpected but easily accessible habitats, such as their private garden, compared to forests or parks (Walter et al., 2018). Therefore, areas with a high report frequency do not necessarily correspond to high species densities, nor do areas without reports mean that the species is absent. Further investigation of such correlations between report numbers and densities is essential to improve the interpretative depth of research with an analogue design to the current study. With this in mind, well-established citizen science monitoring can serve as an important tool for better understanding the dynamics of human–wildlife encounters.
The used R code is available at https://doi.org/10.5281/zenodo.16358732 (Issel, 2025, last access: 2 October 2025).
Data from the platform “Roadkill” are open-source available (https://doi.org/10.15468/ejb47y, BOKU University, Vienna, 2024, last access: 2 October 2025). Data from the platform “StadtWildTiere” and the green space monitoring from the city of Vienna are not publicly available and cannot be shared by the authors for legal reasons.
DD and FH contributed to the research design, the data analysis and the writing of the paper. RZ contributed to the research design and the data management. JGZ contributed to the writing of the paper. JL contributed to the statistical analysis and the writing of the paper. GL contributed to the statistical analysis.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
This research was conducted with the generous support of the citizen science platforms StadtWildTiereWien and Roadkill and the city of Vienna, especially the Municipal Department 22 – Environmental Protection in Vienna.
This paper was edited by Sonja Knapp and reviewed by Michael Strohbach and one anonymous referee.
App, M., Strohbach, M. W., Schneider, A. K., and Schröder, B.: Making the case for gardens: Estimating the contribution of urban gardens to habitat provision and connectivity based on hedgehogs (Erinaceus europaeus), Landscape Urban Plan., 220, 104347, https://doi.org/10.1016/j.landurbplan.2021.104347, 2022.
Araújo, M. B.: The coincidence of people and biodiversity in Europe, Global Ecol. Biogeogr., 12, 5–12, https://doi.org/10.1046/j.1466-822X.2003.00314.x, 2003.
Auguie, B.: gridExtra: Miscellaneous Functions for “Grid” Graphics, R package version 2.3, https://CRAN.R-project.org/package=gridExtra (last access: 26 February 2025), 2017.
Baker, P. J. and Harris, S.: Urban mammals: what does the future hold? An analysis of the factors affecting patterns of use of residential gardens in Great Britain, Mam. Rev., 37, 297–315, https://doi.org/10.1111/j.1365-2907.2007.00102.x, 2007.
Balestrieri, A., Remonti, L., and Prigioni, C.: Habitat Selection in a Low-Density Badger Meles meles Population: A Comparison of Radio-Tracking and Latrine Surveys, Wildlife Biol., 15, 442–448, https://doi.org/10.2981/08-027, 2009.
Balestrieri, A., Cardarelli, E., Pandini, M., Remonti, L., Saino, N., and Prigioni, C.: Spatial organisation of European badger (Meles meles) in northern Italy as assessed by camera-trapping, Eur. J. Wildl. Res., 62, 219–226, https://doi.org/10.1007/s10344-016-0996-y, 2016.
BOKU University, Vienna: Roadkill. Occurrence dataset, GBIF [data set], https://doi.org/10.15468/ejb47y, 2024.
Braaker, S., Kormann, U., Bontadina, F., and Obrist, M. K.: Prediction of genetic connectivity in urban ecosystems by combining detailed movement data, genetic data and multi-path modelling, Landscape Urban Plan., 160, 107–114, https://doi.org/10.1016/j.landurbplan.2016.12.011, 2017.
Davison, J., Huck, M., Delahay, R. J., and Roper, T. J.: Restricted ranging behaviour in a high-density population of urban badgers, J. Zool., 277, 45–53, https://doi.org/10.1111/j.1469-7998.2008.00509.x, 2009.
Doncaster, C. P.: Testing the role of intraguild predation in regulating hedgehog populations, Proceedings of the Royal Society B: Biological Sciences, 249, 113–117, https://doi.org/10.1098/rspb.1992.0092, 1992.
Dowding, C. V., Harris, S., Poulton, S., and Baker, P. J.: Nocturnal ranging behaviour of urban hedgehogs, Erinaceus europaeus, in relation to risk and reward, Anim. Behav., 80, 13–21, https://doi.org/10.1016/j.anbehav.2010.04.007, 2010.
Feore, S. and Montgomery, W. I.: Habitat effects on the spatial ecology of the European badger (Meles meles), J. Zool., 247, 537–549, https://doi.org/10.1111/j.1469-7998.1999.tb01015.x, 1999.
Fischer, C. and Weber, J.-M.: Distribution of badger setts and latrines in an intensively cultivated landscape, Rev. Suisse Zool., 110, 661–668, https://doi.org/10.5962/bhl.part.80204, 2003.
Fox, J. and Weisberg, S.: An R Companion to Applied Regression, Third edition, Sage, Thousand Oaks CA, https://www.john-fox.ca/Companion/ (last access: 26 February 2025), 2019.
Gazzard, A. and Baker, P. J.: Patterns of Feeding by Householders Affect Activity of Hedgehogs (Erinaceus europaeus) during the Hibernation Period, Animals 10, 1344, https://doi.org/10.3390/ani10081344, 2020.
Gazzard, A. and Rasmussen, S. L.: Erinaceus europaeus, The IUCN Red List of Threatened Species 2024, e.T29650A213411773, https://doi.org/10.2305/IUCN.UK.2024-2.RLTS.T29650A213411773.en, (last access: 7 November 2024).
Gazzard, A., Yarnell, R. W., and Baker, P. J.: Fine-scale habitat selection of a small mammalian urban adapter: the West European hedgehog (Erinaceus europaeus), Mamm. Biol., 102, 387–403, https://doi.org/10.1007/s42991-022-00251-5, 2022.
Geiger, M., Taucher, A. L., Gloor, S., Hegglin, D., and Botadina, F.: In the footsteps of city foxes: evidence for a rise of urban badger populations in Switzerland, Hystrix, 236–238, https://doi.org/10.4404/hystrix-00069-2018, 2018.
Goldstein, B. R. and de Valpine, P.: Comparing N−mixture models and GLMMs for relative abundance estimation in a citizen science dataset, Sci Rep 12, 12276, https://doi.org/10.1038/s41598-022-16368-z, 2022.
Gomes, D. J., Wierzbowska, I. A., Bevanger, K., O'Mahony, D. T., and Rola, K.: Diet of the European badgers (Meles meles) in urban and rural areas of Norway, Eur. J. Wildl. Res., 66, https://doi.org/10.1007/s10344-019-1347-6, 2019.
Haberfellner, J., Kutschker, M., Jäpel, J., and Lutter, J.: Wiener Umweltbericht 2020/2021, Stadt Wien-Umweltschutz, 2022.
Harris, S.: Ecology of Urban badgers Meles meles: Distribution in Britain and habitat selection, persecution, food and damage in the city of Bristol, Biol. Conserv., 28, 349–375, https://doi.org/10.1016/0006-3207(84)90041-7, 1984.
Heigl, F., Horvath, K., Laaha, G., Zaller, J.: Amphibian and reptile road-kills on tertiary roads in relation to landscape structure: using a citizen science approach with open-access land cover data, BMC Ecol., 17, 24, https://doi.org/10.1186/s12898-017-0134-z, 2017.
Heigl, F., Teufelbauer, N., Resch, S., Schweiger, S., Stückler, S., and Dörler, D.: A dataset of road-killed vertebrates collected via citizen science from 2014-2020, Sci. Data, 9, 504, https://doi.org/10.1038/s41597-022-01599-6, 2022.
Heikinheimo, V., Tenkanen, H., Bergroth, C., Järv, O., Hiippala, T., and Toivonen, T.: Understanding the use of urban green spaces from user-generated geographic information, Landscape Urban Plan., 201, https://doi.org/10.1016/j.landurbplan.2020.103845, 2020.
Hof, A. R.: A study of the current status of the hedgehog (Erinaceus europaeus), and its decline in Great Britain since 1960, PhD thesis, Roywal Holloway University of London, 2009.
Hof, A. R. and Bright, P. W.: The value of green-spaces in built-up areas for hedgehogs, Lutra, 69–82, 2009.
Hof, A. R., Snellenberg, J., and Bright, P. W.: Food or fear? Predation risk mediates edge refuging in an insectivorous mammal, Anim. Behav., 83, 1099–1106, https://doi.org/10.1016/j.anbehav.2012.01.042, 2012.
Hof, A. R., Allen, A. M., and Bright, P. W.: Investigating the Role of the Eurasian Badger (Meles meles) in the Nationwide Distribution of the Western European Hedgehog (Erinaceus europaeus) in England, Animals, 9, https://doi.org/10.3390/ani9100759, 2019.
Hubert, P., Julliard, R., Biagianti, S., and Poulle, M.-L.: Ecological factors driving the higher hedgehog (Erinaceus europeaus) density in an urban area compared to the adjacent rural area, Landscape Urban Plan., 103, 34–43, https://doi.org/10.1016/j.landurbplan.2011.05.010, 2011.
Huck, M., Davison, J., and Roper, T. J.: Predicting European badger Meles meles sett distribution in urban environments, Wildlife Biol., 14, 188–198, https://doi.org/10.2981/0909-6396(2008)14[188:PEBMMS]2.0.CO;2, 2008.
Issel, D.: Urban wildlife monitoring using citizen science suggests that European hedgehogs and badgers select different habitats, Zenodo [code], https://doi.org/10.5281/zenodo.16358732, 2025.
Ives, C. D., Lentini, P. E., Threlfall, C. G., Ikin, K., Shanahan, D. F., Garrard, G. E., Bekessy, S. A., Fuller, R. A., Mumaw, L., Rayner, L., Rowe, R., Valentine, L. E., and Kendal, D.: Cities are hotspots for threatened species, Global Ecol. Biogeogr., 25, 117–126, https://doi.org/10.1111/geb.12404, 2016.
Kettel, E. F., Gentle, L. K., Quinn, J. L., and Yarnell, R. W.: The breeding performance of raptors in urban landscapes: a review and meta-analysis, J. Ornithol., 159, 1–18, https://doi.org/10.1007/s10336-017-1497-9, 2018.
Korslund, L. M., Floden, M. S., Albertsen, M. M. S., Landsverk, A., Løkken, K. M. V., and Johansen, B. S.: Home Range, Movement, and Nest Use of Hedgehogs (Erinaceus europaeus) in an Urban Environment Prior to Hibernation, Animals, 14, https://doi.org/10.3390/ani14010130, 2023.
Kranz, A., Abramov, A. V., Herrero, J., and Maran, T.: Meles meles, The IUCN Red List of Threatened Species 2016, https://doi.org/10.2305/IUCN.UK.2016-1.RLTS.T29673A45203002.en, 2016.
Kruuk, H. and Parish, T.: Feeding Specialization of the European Badger Meles meles in Scotland, J. Anim. Ecol., 50, 773–788, https://doi.org/10.2307/4136, 1981.
Kutner, M. H., Nachtsheim, C., and Neter, J.: Applied linear regression models, 4th edn., The McGraw-Hill/Irwin series: operations and decision sciences, 1 online resource, McGraw-Hill/Irwin, Boston, New York, ISBN 0072386916, 2004.
Levandowsky, M. and Winter, D.: Distance between Sets, Nature, 234, 34–35, https://doi.org/10.1038/234034a0, 1971.
Łopucki, R., Kitowski, I., Perlińska-Teresiak, M., and Klich, D.: How Is Wildlife Affected by the COVID-19 Pandemic? Lockdown Effect on the Road Mortality of Hedgehogs, Animals, 11, https://doi.org/10.3390/ani11030868, 2021.
Lovell, C., Li, S., Turner, J., and Carbone, C.: The effect of habitat and human disturbance on the spatiotemporal activity of two urban carnivores: The results of an intensive camera trap study, Ecol. Evol., 12, e8746, https://doi.org/10.1002/ece3.8746, 2022.
Lowry, H., Lill, A., and Wong, B.: Behavioural responses of wildlife to urban environments, Biol. Rev., 88, https://doi.org/10.1111/brv.12012, 2013.
Lumley, T., Diehr, P., Emerson, S., and Chen, L.: The importance of the normality assumption in large public health data sets, Annu. Rev. Publ. Health, 23, 151–169, https://doi.org/10.1146/annurev.publhealth.23.100901.140546, 2002.
Macdonald, D. W., Newman, C., Dean, J., Buesching, C. D., and Johnson, P. J.: The distribution of Eurasian badger, Meles meles, setts in a high-density area: field observations contradict the sett dispersion hypothesis, Oikos, 106, 295–307, https://doi.org/10.1111/j.0030-1299.2004.12879.x, 2004.
Marco-Tresserras, J. and López-Iborra, G. M.: The effect of sex on home range in an urban population of European hedgehogs Erinaceus europaeus at the southern edge of its distribution, Anim. Biodiv. Conserv., 269–279, https://doi.org/10.32800/abc.2022.45.0269, 2022.
Matthews, F., Kubasiewicz, L. M., Gurnell, J., Harrower, C. A., McDonald, R. A., and Shore, R. F.: A review of the population and conservation status of British mammals, Natural England Joint Publication, JP025, Natural England, Peterborough, England, ISBN 978-1-78354-494-3, 699 pp., 2018.
Micol, T., Doncaster, C. P., and Mackinlay, L. A.: Correlates of Local Variation in the Abundance of Hedgehogs Erinaceus europaeus, J. Anim. Ecol., 63, 851, https://doi.org/10.2307/5262, 1994.
Microsoft Corporation: Microsoft® Excel® for Microsoft 365 MSO (Version 2501 Build 16.0.18429.20132) 64 Bit, https://www.microsoft.com/en-us/microsoft-365/excel (last access: 10 October 2024).
Morris, P. A.: A study of home range and movements in the hedgehog (Erinaceus europaeus), J. Zool., 214, 433–449, https://doi.org/10.1111/j.1469-7998.1988.tb03751.x, 1988.
Nottingham, C., Glen, A., and Stanley, M.: Snacks in the city: the diet of hedgehogs in Auckland urban forest fragments, New Zeal. J. Ecol., 43, https://doi.org/10.20417/nzjecol.43.24, 2019.
Obidziński, A., Pabjanek, P., and Mędrzycki, P.: Determinants of badger Meles meles sett location in Białowieża Primeval Forest, northeastern Poland, Wildlife Biol., 19, 48–68, https://doi.org/10.2981/11-074, 2013.
Pettett, C. E., Moorhouse, T. P., Johnson, P. J., and Macdonald, D. W.: Factors affecting hedgehog (Erinaceus europaeus) attraction to rural villages in arable landscapes, Eur. J. Wildl. Res., 63, https://doi.org/10.1007/s10344-017-1113-6, 2017.
Pettett, C. E., Johnson, P. J., Moorhouse, T. P., and Macdonald, D. W.: National predictors of hedgehog Erinaceus europaeus distribution and decline in Britain, Mamm. Rev., 48, 1–6, https://doi.org/10.1111/mam.12107, 2018.
Piza-Roca, C., van 't Zelfde, M., La Haye, M. J. J., Jongejans, E., and Raes, N.: European badger habitat requirements in the Netherlands – combining ecological niche models with neighbourhood analysis, Wildlife Biol., 2018, 1–11, https://doi.org/10.2981/wlb.00453, 2018.
QGIS Development Team: QGIS Geographic Information System, Open Source Geospatial Foundation Project, https://qgis.org/ (last access: 26 February 2025), 2023.
Rasmussen, S. L., Berg, T. B., Martens, H. J., and Jones, O. R.: Anyone Can Get Old-All You Have to Do Is Live Long Enough: Understanding Mortality and Life Expectancy in European Hedgehogs (Erinaceus europaeus), Animals, 13, https://doi.org/10.3390/ani13040626, 2023.
Rautio, A., Isomursu, M., Valtonen, A., Hirvelä-Koski, V., and Kunnasranta, M.: Mortality, diseases and diet of European hedgehogs (Erinaceus europaeus) in an urban environment in Finland, Mamm. Res., 61, 161–169, https://doi.org/10.1007/s13364-015-0256-7, 2015.
R Core Team: R: A Language and Environment for Statistical Computing, R Foundation for Statistical Computing, Vienna, Austria, https://www.R-project.org/ (last access: 26 February 2025), 2023.
Reid, N., Etherington, T. R., Wilson, G. J., Montgomery, W. I., and McDonald, R. A.: Monitoring and population estimation of the European badger Meles meles in Northern Ireland, Wildlife Biol., 18, 46–57, https://doi.org/10.2981/11-016, 2012.
Rondinini, C. and Doncaster, C. P.: Roads as barriers to movement for hedgehogs, Funct. Ecol., 16, 504–509, https://doi.org/10.1046/j.1365-2435.2002.00651.x, 2002.
RStudio Team: RStudio: Integrated Development Environment for R (Version 2024.04.2 Build 764), RStudio, PBC, https://posit.co (last access: 26 February 2025), 2024.
Šálek, M., Drahníková, L., and Tkadlec, E.: Changes in home range sizes and population densities of carnivore species along the natural to urban habitat gradient, Mamm. Rev., 45, 1–14, https://doi.org/10.1111/mam.12027, 2015.
Santini, L., González-Suárez, M., Russo, D., Gonzalez-Voyer, A., Von Hardenberg, A., and Ancillotto, L.: One strategy does not fit all: determinants of urban adaptation in mammals, Ecol. Lett., 22, https://doi.org/10.1111/ele.13199, 2018.
Schaus, J., Uzal, A., Gentle, L. K., Baker, P. J., Bearman-Brown, L., Bullion, S., Gazzard, A., Lockwood, H., North, A., Reader, T., Scott, D. M., Sutherland, C. S., and Yarnell, R. W.: Application of the Random Encounter Model in citizen science projects to monitor animal densities, Remote Sens. Ecol. Conserv., 6, 514–528, https://doi.org/10.1002/rse2.153, 2020.
Schley, L., Schaul, M., and Ropert, T. J.: Distribution and population density of badgers Meles meles in Luxembourg, Mam. Rev., 34, 233–240, https://doi.org/10.1111/j.1365-2907.2004.00040.x, 2004.
Schwierz, G. and Wachendörfer, G.: Studie über die Ursachen des starken Rückganges der Dachspopulation in Hessen im Zeitraum 1952–1977, Eur. J. Wildl. Res., 27, 145–153, https://doi.org/10.1007/BF02243708, 1981.
Smith, G. and Wilkinson, D.: Modelling disease spread in a novel host: rabies in the European badger Meles meles. Journal of Applied Ecology, 39, 865–874, https://doi.org/10.1046/j.1365-2664.2002.00773.x, 2002.
Taucher, A. L., Gloor, S., Dietrich, A., Geiger, M., Hegglin, D., and Bontadina, F.: Decline in Distribution and Abundance: Urban Hedgehogs under Pressure, Animals, 10, https://doi.org/10.3390/ani10091606, 2020.
Tiago, P., Ceia-Hasse, A., Marques, T. A., Capinha, C. and Pereira H. M.: Spatial distribution of citizen science casuistic observations for different taxonomic groups, Sci Rep 7, 12832, https://doi.org/10.1038/s41598-017-13130-8, 2017.
United Nations, Department of Economic and Social Affairs, Population Division (UNDESA): World urbanization prospects: The 2018 revision, New York, 103 pp., 2019.
University of Natural Resources and Life Sciences, Vienna: Roadkill, Occurrence dataset, GBIF [data set], https://doi.org/10.15468/ejb47y, 2024.
van de Poel, J. L., Dekker, J., and Langevelde, F.: Dutch hedgehogs Erinaceus europaeus are nowadays mainly found in urban areas, possibly due to the negative Effects of badgers Meles meles, Wildlife Biol., 21, 51–55, https://doi.org/10.2981/wlb.00072, 2015.
van den Berge, K., van der Veken, T., Gouwy, J., Verschelde, P., and Eeraerts, M.: Dietary composition and overlap among small- and medium-sized carnivores in Flanders, Belgium, Ecol. Res., 37, 163–170, https://doi.org/10.1111/1440-1703.12276, 2022.
Venables, W. N. and Ripley, B. D.: Modern Applied Statistics with S, Fourth Edition, Springer, New York, ISBN 0-387-95457-0, 2002.
Walter, T., Zink, R., Laaha, G., Zaller, J. G., and Heigl, F.: Fox sightings in a city are related to certain land use classes and sociodemographics: results from a citizen science project, BMC Ecol., 18, 50, https://doi.org/10.1186/s12898-018-0207-7, 2018.
Wickham, H.: ggplot2: Elegant Graphics for Data Analysis, Springer-Verlag New York, https://doi.org/10.1007/978-3-319-24277-4, 2016.
Wickham, H., Averick, M., Bryan, J., Chang, W., McGowan, L. D., François, R., Grolemund, G., Hayes, A., Henry, L., Hester, J., Kuhn, M., Pedersen, T. L., Miller, E., Bache, S. M., Müller, K., Ooms, J., Robinson, D., Seidel, D. P., Spinu, V., Takahashi, K., Vaughan, D., Wilke, C., Woo, K., and Yutani, H.: “Welcome to the tidyverse.”, Journal of Open Source Software, 4, 1686, https://doi.org/10.21105/joss.01686, 2019.
Williams, B. M., Baker, P. J., Thomas, E., Wilson, G., Judge, J., and Yarnell, R. W.: Reduced occupancy of hedgehogs (Erinaceus europaeus) in rural England and Wales: The influence of habitat and an asymmetric intra-guild predator, Sci. Rep.-UK, 8, 12156, https://doi.org/10.1038/s41598-018-30130-4, 2018a.
Williams, B., Mann, N., Neumann, J. L., Yarnell, R. W., and Baker, P. J.: A prickly problem: developing a volunteer-friendly tool for monitoring populations of a terrestrial urban mammal, the West European hedgehog (Erinaceus europaeus), Urban Ecosyst., 21, 1075–1086, https://doi.org/10.1007/s11252-018-0795-1, 2018b.
Williams, R. L., Stafford, R., and Goodenough, A. E.: Biodiversity in urban gardens: Assessing the accuracy of citizen science data on garden hedgehogs, Urban Ecosyst., 18, 819–833, https://doi.org/10.1007/s11252-014-0431-7, 2015.
Wine, S., Gagné, S. A., and Meentemeyer, R. K.: Understanding human–coyote encounters in urban ecosystems using citizen science data: what do socioeconomics tell us?, Environ. Manage., 55, 159–170, https://doi.org/10.1007/s00267-014-0373-0, 2015.
Wright, P. G. R., Coomber, F. G., Bellamy, C. C., Perkins, S. E., and Mathews, F.: Predicting hedgehog mortality risks on British roads using habitat suitability modelling, PeerJ, 7, e8154, https://doi.org/10.7717/peerj.8154, 2020.
Young, R. P., Davison, J., Trewby, I. D., Wilson, G. J., Delahay, R. J., and Doncaster, C. P.: Abundance of hedgehogs (Erinaceus europaeus) in relation to the density and distribution of badgers (Meles meles), J. Zool., 269, 349–356, https://doi.org/10.1111/j.1469-7998.2006.00078.x, 2006.
Zink, R. and Walter, T.: Where pathways cross: citizen science project StadtWildTiere in Vienna, Austria, Front. Environ. Sci. Conference Abstract, Austrian Citizen Science Conference 2016, https://doi.org/10.3389/conf.FENVS.2016.01.00007, 2016.