the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 26 May 2026
| 26 May 2026
Intraspecific neighbourhood effects on male and female fitness in Moricandia moricandioides
Violeta Quiroga-Álvarez
Cristina Poyatos
Inés Álvarez
Luis G. Quintanilla
Luis Giménez-Benavides
A. Jesús Muñoz-Pajares
Rubén Torices
The spatial arrangement of individuals within plant populations is largely shaped by seed dispersal mechanisms, resulting in characteristic patterns of individual and genetic aggregation. These configurations can directly influence individual fitness, especially in insect-pollinated and self-incompatible species, where neighbourhood density and genetic relatedness may influence pollinator visitation and the likelihood of compatible mating events. In this study, we analysed a natural population of Moricandia moricandioides, a hermaphroditic, self-incompatible, and insect-pollinated species. All individuals in a 15 m×15 m plot were mapped and sampled for genetic analyses. We quantified floral traits (flower number and corolla diameter), fruit, and seed production in 50 of the 179 plants in the plot. Male reproductive success was estimated through the paternity analyses of seedlings. We detected a pattern of isolation by distance. The presence of neighbouring plants was associated with a reduced fruit production, while floral traits significantly influenced all components of female reproductive success, independently of neighbourhood context. In contrast, siring success depended on the interaction between floral traits and the presence of conspecific neighbours. Furthermore, the number of neighbours and their genetic relatedness had a significant negative effect on siring success. However, they were associated with an increase in the mean genetic distance to pollen donors, suggesting that local genetic structure can influence mating patterns. These findings illustrate the complex interplay among neighbourhood composition, floral display, and reproductive success in M. moricandioides, contributing to our understanding of how ecological and genetic neighbourhoods shape plant reproductive strategies.
- Article
(2028 KB) - Full-text XML
-
Supplement
(409 KB) - BibTeX
- EndNote




The spatial arrangement of individuals within plant populations is rarely random; rather, it exhibits spatial structuring (Epperson, 1992; Escudero et al., 2003; Loveless and Hamrick, 1984; Vekemans and Hardy, 2004). This spatial structure is mostly determined both by microhabitat availability and the plant dispersal capacity (Nogueira et al., 2014; Schupp et al., 2010). Within populations, individual positioning is largely shaped by seed dispersal mechanisms, resulting in characteristic patterns of spatial aggregation (Nathan and Muller-Landau, 2000; Peakall and Beattie, 1995). A common outcome is the clumping of offspring around maternal plants due to limited dispersal distances, often resulting in fine-scale genetic structuring (e.g. Hardy et al., 2006; Jones and Hubbell, 2006). Such spatial genetic configurations can have direct consequences for individual fitness and population dynamics.
In pollinator-mediated species, the spatial aggregation of plants can significantly influence pollinator foraging behaviour. Pollinators tend to concentrate their activity in areas with high floral density, as this maximizes their energy intake per unit effort – a pattern consistent with predictions from optimal foraging theory (Biernaskie et al., 2009; Hegland, 2014; Moeller, 2004; Thomson, 1981). Consequently, the local neighbourhood context of a plant may affect pollinator visitation rates, thereby shaping mating patterns and ultimately influencing reproductive success (Dauber et al., 2010; Grindeland et al., 2005; Klinkhamer et al., 2001; Knight, 2003; Martén-Rodríguez et al., 2012). In hermaphroditic outcrossing species, the net effect of conspecific aggregation on pollinator behaviour may have different consequences for male and female reproductive functions at the level of individual plants, which could result in differences in reproductive success among neighbouring individuals.
The interaction between conspecific spatial aggregation and pollinator behaviour can promote non-random mating among genetically related individuals. Since individuals located in close spatial proximity are expected to exhibit a higher degree of genetic similarity than expected by chance, this would lead to an increase in mating events between genetically related individuals (Dutech et al., 2005; Vekemans and Hardy, 2004). Thus, in pollinator-mediated outcrossing species, limited pollen dispersal and neighbourhood structure can lead to genetic structuring at fine spatial scales, which might impose fitness costs through increased mating among close relatives (Charlesworth and Willis, 2009; Hardy et al., 2006; Stacy et al., 1996). Consequently, genetic homogeneity within population patches would also increase, thereby enhancing the expression of deleterious alleles and ultimately reducing offspring fitness (Charlesworth and Charlesworth, 1987; Charlesworth and Willis, 2009). However, conspecific aggregation can also enhance reproductive success through increased pollinator attraction and pollen export. Neighbourhoods with higher plant density often act as floral magnets, attracting more pollinators and thereby increasing the probability of pollen transfer to conspecifics (Ghazoul, 2005; Hurtado et al., 2023; Underwood et al., 2020). This magnet effect may facilitate pollinator sharing among neighbouring plants and increase opportunities for pollen export. As a result, individual plants may increase their relative male reproductive success by siring a larger proportion of the seeds in the patch (Laverty, 1992; Ruttan et al., 2021). In Moricandia moricandioides (Boiss.) – a short-lived self-incompatible herb – floral display has been shown to be significantly enhanced both in terms of flower number and petal biomass when individuals grow among conspecifics, particularly among genetically related neighbours (Torices et al., 2018). This suggests a potential cooperative strategy, where kin groups invest more in floral advertising, which might have been selected by indirect fitness benefits derived from the reproductive success of relatives (Sun et al., 2021). However, the combined effects of neighbourhood density and genetic relatedness on male and female fitness components have received little empirical attention in natural populations.
In patchy environments, understanding how spatial and genetic neighbourhoods influence male and female components of fitness can elucidate the evolutionary dynamics of plant reproductive strategies. Local neighbourhoods can have contrasting effects on different components of plant fitness, potentially promoting both cooperation and conflict among conspecifics. Based on previous evidence of enhanced floral display in aggregated kin neighbourhoods, we hypothesized that the neighbourhood context may influence plant reproductive success through its effects on pollinator attraction and mating opportunities. Specifically, higher conspecific density is expected to increase pollinator visitation rates and opportunities for pollen export, potentially enhancing male reproductive success. In contrast, high genetic relatedness among neighbouring plants may impose compatibility constraints in this self-incompatible species, limiting the female reproductive success. Although floral traits such as flower number and corolla size influence pollinator attraction, these traits may have different fitness consequences for solitary and aggregated plants. For instance, these floral display traits may affect pollinator foraging behaviour at two key stages in the decision-making process: first, selecting which patches to visit; and second, choosing which flowers in a patch to visit.
In this study, we aimed to investigate how neighbourhood composition influences male and female reproductive success in M. moricandioides. Specifically, we aimed to (i) characterize the fine-scale genetic structure of a natural population; and (ii) evaluate how neighbourhood effects – considering both conspecific density and genetic relatedness among neighbouring plants – influence male and female fitness components, and how the neighbourhood may influence the relationship between floral display and both fitness components.
2.1 Study species and location
Moricandia moricandioides (Boiss.) Heywood (Brassicaceae) is a predominantly annual, hermaphroditic, and self-incompatible species that inhabits semi-arid environments in southeastern and central Spain, including gypsum-rich soils. Occasionally it overwinters as a vegetative leaf rosette and initiates flowering in April and May (Gómez, 1996; Sobrino Vesperinas, 1993), with fruit development occurring in June. The species is insect pollinated, primarily by long-tongued bees, notably those from the genera Anthophora and Eucera (Gómez, 1996; Gómez et al., 2016). Seeds disperse when mature, and dry fruits open and release seeds in close proximity to the parent plants.
The study was conducted in a natural population of M. moricandioides at Soto de Cerrato (Spain: N, W; elevation: 873–880 m a.s.l.). The study location has a temperate climate with dry summers, characterized by a mean annual temperature of 11.3 °C and an average yearly precipitation of 504 mm (Agroclimatic Atlas of the Technological Agricultural Institute of Castilla y León and the State Meteorological Agency: http://www.atlas.itacyl.es/, last access: 28 October 2025). The soils are lithic leptosols and calcareous regosols developed over basic sedimentary rocks, primarily limestone and marl (Agroclimatic Atlas of the Technological Agricultural Institute of Castilla y León and the State Meteorological Agency: http://www.atlas.itacyl.es/, last access: 28 October 2025). The dominant vegetation consists of grassland and low-growing shrubland with a prevalence of Chamaephytes such as Hedysarum boveanum subsp. palentinum Valdés, Helianthemum spp., Linum suffruticosum L., and Lithodora fruticosa Griseb., interspersed with pine plantations (Pinus spp.) and deciduous broad-leaved trees like gall oaks (Quercus faginea).
2.2 Mapping of individuals and spatial structure
The population was sampled in May 2022 by establishing a 15 m×15 m plot. Using a differential GPS (Leica Viva GNSS GPS GS15_1500336), all 179 individuals present in the plot were mapped. Plants within the plot showed a spatial distribution consistent with an inhomogeneous Poisson cluster process (Quiroga-Álvarez et al., unpublished). This indicates that individuals were significantly aggregated, but the intensity of aggregation varied heterogeneously across space. The average radius of the aggregates predicted by this model was 0.35 m (Quiroga-Álvarez et al., unpublished).
Fresh leaves were collected from all individuals within the plot for subsequent genetic analyses including DNA extraction and sequencing. Additionally, to characterize floral traits, and assess fruit and seed production, 50 individuals were sampled across the plot. To this end, the plot was divided into 25 square cells, each measuring 3 m per side. In each cell, two flowering individuals were randomly selected and tagged. For each of the 50 tagged plants, we measured the maximum corolla diameter in two to three flowers, recorded the total number of flowers and fruits, and counted seeds in 10 unripened fruits. The total seed number per individual was estimated by multiplying the mean number of seeds per fruit by the total number of fruits produced. Fruit set was calculated as the proportion of flowers that developed into fruits. In this species, the total number of flowers can be estimated at the end of the flowering season because all flowers leave a visible mark on the stem, regardless of whether they develop into fruit. Therefore, all flowers and fruits produced by a single plant can be estimated at fruiting stage.
2.3 Genomic data processing and filtering
DNA extractions were performed from the fresh material collected in the field using the CTAB-based protocol (Doyle and Doyle, 1987) modified for suboptimal samples (Larridon et al., 2020) and were carried out at the Real Jardín Botánico, CSIC (Spain). Subsequently we prepared the genomic libraries using the kit Collibri™ ES DNA Library Prep Kit for Illumina Systems at the University of Granada (Spain). The resulting libraries were sequenced by Novogene Inc. using two lanes of the HiSeq × System (Illumina) at low coverage (5X). Raw reads for each individual were mapped to a reference genome available on NCBI (M. moricandioides, accession: GCA_905132885.1) using BWA-MEM (Li, 2013). For each sample, alignments were sorted and converted to BAM format with SAMtools (Li et al., 2009). For each BAM file, we generated binary variant call files (BCFs) using BCFtools (Danecek et al., 2021). The resulting BCF files were merged for downstream filtering. Single nucleotide polymorphisms (SNPs) were subsequently filtered using VCFtools (Danecek et al., 2011) and PLINK v1.9 (Purcell et al., 2007), retaining only biallelic variants that met specific thresholds for quality (QUAL>20), read missingness (m<15), and linkage disequilibrium (r2<0.1). This filtering resulted in a total of 90 337 SNPs across 179 individuals.
2.4 Paternity assignments and male fitness estimation
Male reproductive success was quantified through paternity analysis based on offspring genotyping. In 2023, seeds from all 50 tagged plants were sown and cultivated at the CULTIVE facilities of Universidad Rey Juan Carlos (Spain). Leaf tissue was sampled from 315 seedlings for genotyping. However, due to logistical constraints imposed by concurrent experiments, the final number of genotyped offspring varied among maternal plants (Fig. 1). We followed the same protocol described above, except for DNA extraction, which was performed using the NZY Plant/Fungi gDNA Isolation kit (NZYTech, Lisbon, Portugal; cat. no. MB17702). Prior to analysis, SNP data were filtered to retain only loci with a minor allele frequency greater than 0.3. Paternity analyses were conducted using the Sequoia R package (Huisman, 2017), assuming hermaphroditism and discrete generations. A high threshold of 500 was set for the maximum sibship size to avoid artificially restricting parental assignments in the population, and a constraint matrix was built to incorporate known maternal relationships. A total of 270 paternity relationships were correctly assigned to a most-likely father, while the remaining 45 seedlings (14.28 %) could not be confidently assigned to any candidate father in the population.
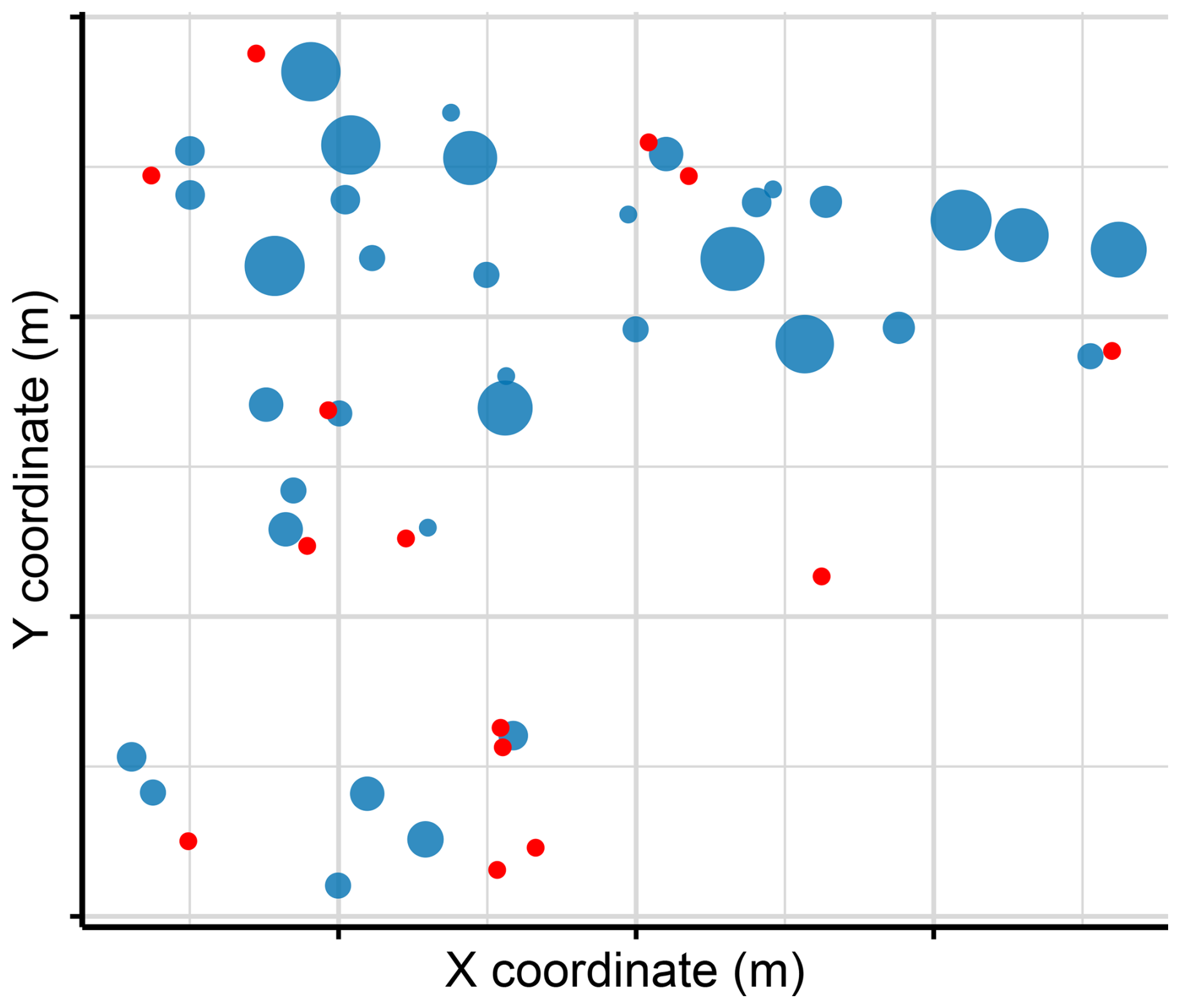
Figure 1Spatial distribution of maternal plants within the study plot showing the number of genotyped offspring per mother. Blue circles represent mothers with at least one genotyped offspring, with point size proportional to the number of offspring. Red points correspond to maternal plants for which no offspring were genotyped.
2.5 Fine-scale genetic structure
To characterize the fine-scale genetic structure (FSGS), we performed an autocorrelation approach implemented in the program SPAGeDI 1.5 (Hardy and Vekemans, 2002). We estimated pairwise genetic relatedness using Loiselle's kinship coefficient Fij (Loiselle et al., 1995) among all individuals in the plot. The slope of the regression between relatedness values and the logarithm of the spatial distances, denoted as bLd, was used to quantify the strength of spatial genetic structure. Standard errors for Fij and bd were estimated by jackknife resampling across loci. To assess the significance of FSGS, we generated a null distribution by randomly permuting genotypes across spatial locations and loci 1000 times.
2.6 Statistical analyses
We evaluated the influence of local neighbourhood conditions on female and male fitness components using two complementary analytical approaches. Female fitness was assessed through fruit set (the proportion of flowers setting fruits), the number of fruits, and the number of seeds, as these metrics are commonly used proxies that reflect successful fertilization events and the number of potential offspring produced by each maternal plant. Male fitness was estimated as siring success, calculated as the proportion of genotyped offspring sired by each pollen donor relative to the total number of genotyped offspring in the population. This estimate was based on paternity analyses of individuals within the plot and seeds collected from fruits of the same population. This proportional metric was used because not all offspring produced in the population were counted or genotyped, thus the total number of seeds sired by each individual could not be determined. The proportion of sired offspring therefore provide a standardized estimate of relative male reproductive success.
In the first approach, we tested whether the presence of conspecifics in the immediate neighbourhood modulated the effect of individual mean floral display traits (corolla diameter and number of flowers) on fitness outcomes. The local neighbourhood was defined as a 0.35 m radius, corresponding to the average size of plant aggregations estimated via spatial point pattern analysis (Quiroga-Álvarez et al., unpublished). We fitted separate generalized linear models (GLMs) using the lme4 package (Bates et al., 2015) implemented in R (version 4.5.1, R Core Team, 2026) for each fitness component. Model assumptions were evaluated using the DHARMa package (Hartig, 2022) to check residual distribution and to detect potential deviations from homoscedasticity or overdispersion. Estimated marginal means and pairwise comparisons were obtained using the emmeans package (Lenth, 2024), and model predictions and graphical representations were produced with ggeffects and ggplot2 (Wickham, 2016; Lüdecke et al., 2023). Explanatory variables included mean floral traits (corolla diameter and number of flowers), conspecific presence within the local neighbourhood, and the interactions between neighbourhood and each floral trait. All explanatory variables were treated as fixed effects. Fruit set and siring success were modelled with a binomial distribution. Fruit and seed counts were modelled with a negative binomial distribution to account for overdispersion in the count data.
In the second approach, we examined how the density and genetic relatedness of conspecifics in the immediate neighbourhood influenced fitness components. This analysis was restricted to individuals surrounded by at least one conspecific within the 0.35 m radius neighbourhood. Separate GLMs were fitted for each fitness component, with the number of neighbouring conspecifics, the average genetic relatedness among them, and their interaction included as explanatory variables using the glmmTMB package in R (Brooks et al., 2017). Fruit set and siring success were modelled using a binomial distribution, whereas fruit and seed counts were fitted using gamma distributions. For seed and fruit counts, we compared models fitted with either binomial negative or gamma distribution. The latter provided a better fit for seed number and performed similarly for fruit number. The model fit was compared using AIC, and model assumptions were evaluated using simulated residual diagnostics implemented in the DHARMa package. Genetic relatedness was estimated using Queller and Goodnight's r coefficient (Queller and Goodnight, 1989), calculated with SPAGeDi 1.5 (Hardy and Vekemans, 2002). This widely used estimator provides pairwise estimates of relatedness, standardized relative to the average relatedness of the population.
To account for potential spatial autocorrelation, we performed Moran's I tests on all response variables using the package ape (Paradis and Schliep, 2019). For the female fitness components, only fruit set exhibited significant spatial autocorrelation (I=0.075, P=0.038), while the other components showed no significant spatial structure (number of fruits: I=0.012, P=0.350; number of seeds: , P=0.655). For the male component, Moran's I test was first calculated using the subset of the 50 marked individuals, which showed no spatial autocorrelation (, P=0.919). We then repeated the analysis using siring success estimated for the entire population, which revealed significant spatial autocorrelation (I=0.042, P=0.009). Accordingly, spatial coordinates were included as a random effect in models where fruit set was the response variable, as well as in the model for the siring success for the entire population.
To assess the effects of the local neighbourhood conditions and floral traits on mating patterns, we fitted GLMs like those described above but using mean mating distance to either the mother or the father as a response variable. First, we tested whether corolla diameter, flower number, and their respective interactions with the presence of conspecifics in the local neighbourhood influenced the mating distance to fathers, assuming a Gaussian distribution, after applying a logarithmic transformation, with the pollen receptor identity used as a random factor. Due to the limited sample size for mating distance from fathers to mothers and for paternal floral display traits, it was not possible to fit a statistical model.
Using the same approach, we also assessed the density and genetic relatedness of neighbouring conspecifics on the mean mating distance to either the mother or the father (see Fig. S1 in the Supplement). We assumed a Gaussian distribution for mean mating distance to the mother and the father after applying a logarithmic transformation and used the pollen receptor and pollen donor identity as a random factor, respectively. Lastly, we tested whether the observed distribution of distances between all mating pairs was significantly different from a null distribution. To build this null distribution, we permuted father identities among mothers 10 000 times while keeping the plant spatial position fixed and recalculated mating distance in each permutation.
3.1 Characterization of fine-scale genetic structure
Mean kinship coefficients (Fij) significantly decreased with the spatial distance between individuals (n=179, ± 0.0007 SE, P<0.001, Fig. 2), indicating a decrease in genetic relatedness of plants as spatial distance within the plot increases. This result is consistent with the expectations under the isolation by distance (IBD) hypothesis, where nearby individuals tend to be more genetically similar than those farther apart. However, only those plants growing at distances below the 0.5 m threshold showed significantly higher kinship coefficients than expected by chance (Fig. 2).
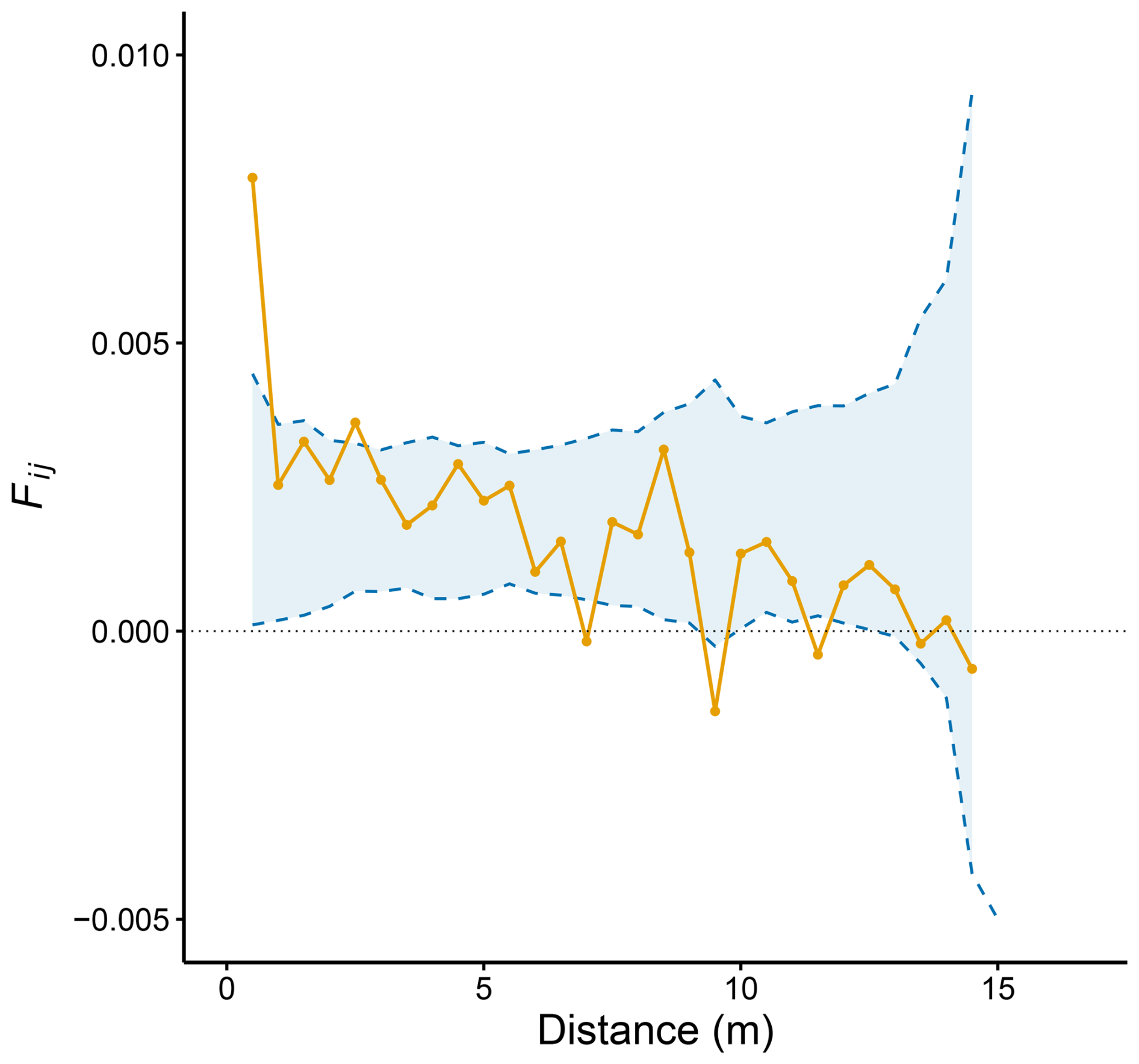
Figure 2Fine-scale spatial genetic structure as measured by pairwise Loiselle's kinship coefficient (Fij), plotted against geographic distance between individuals. The yellow line shows the observed kinship values, while the dashed blue lines and the shaded ribbon indicate the 95 % confidence interval obtained from permutation tests.
3.2 Neighbourhood effects on female and male fitness components
3.2.1 The effect of neighbouring plants on the relationship between floral display and fitness
The presence of neighbouring plants was significantly associated with a reduced fruit production and marginally with smaller fruit set as well as fewer seeds (Table 1). Moreover, individual floral display traits, including both flower number and corolla diameter, significantly influenced all the components of female reproductive success (Table 1). Plants with larger corollas produced a significantly higher number of fruits and seeds as well as higher fruit set (Table 1, Fig. 3a–c). A higher number of flowers also led to a higher number of fruits and seeds but a lower probability of setting fruits (Table 1, Fig. 4). The interaction of neighbourhood with floral display traits did not significantly affect any component of female reproductive success (Table 1).
Table 1The effect of conspecific plant presence in the local neighbourhood, floral display traits (no. of flowers and corolla diameter), and their interaction on plant fitness and mean mating distance. The main effects were assessed using type III effects when the interaction was statistically significant, otherwise type II were used. Models were fitted independently for each one of the four response variables. The spatial coordinates were included as a random effect for the fruit set model. Variance and SD is shown for this random effect. Significant P values (<0.05) are highlighted in bold.
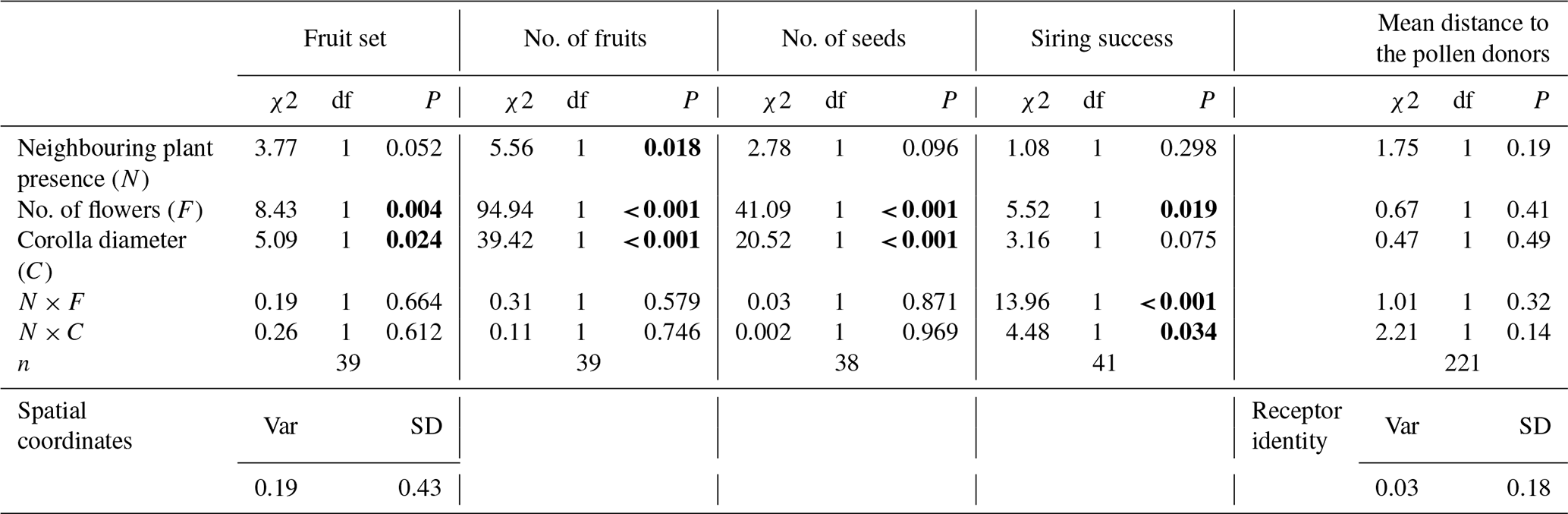
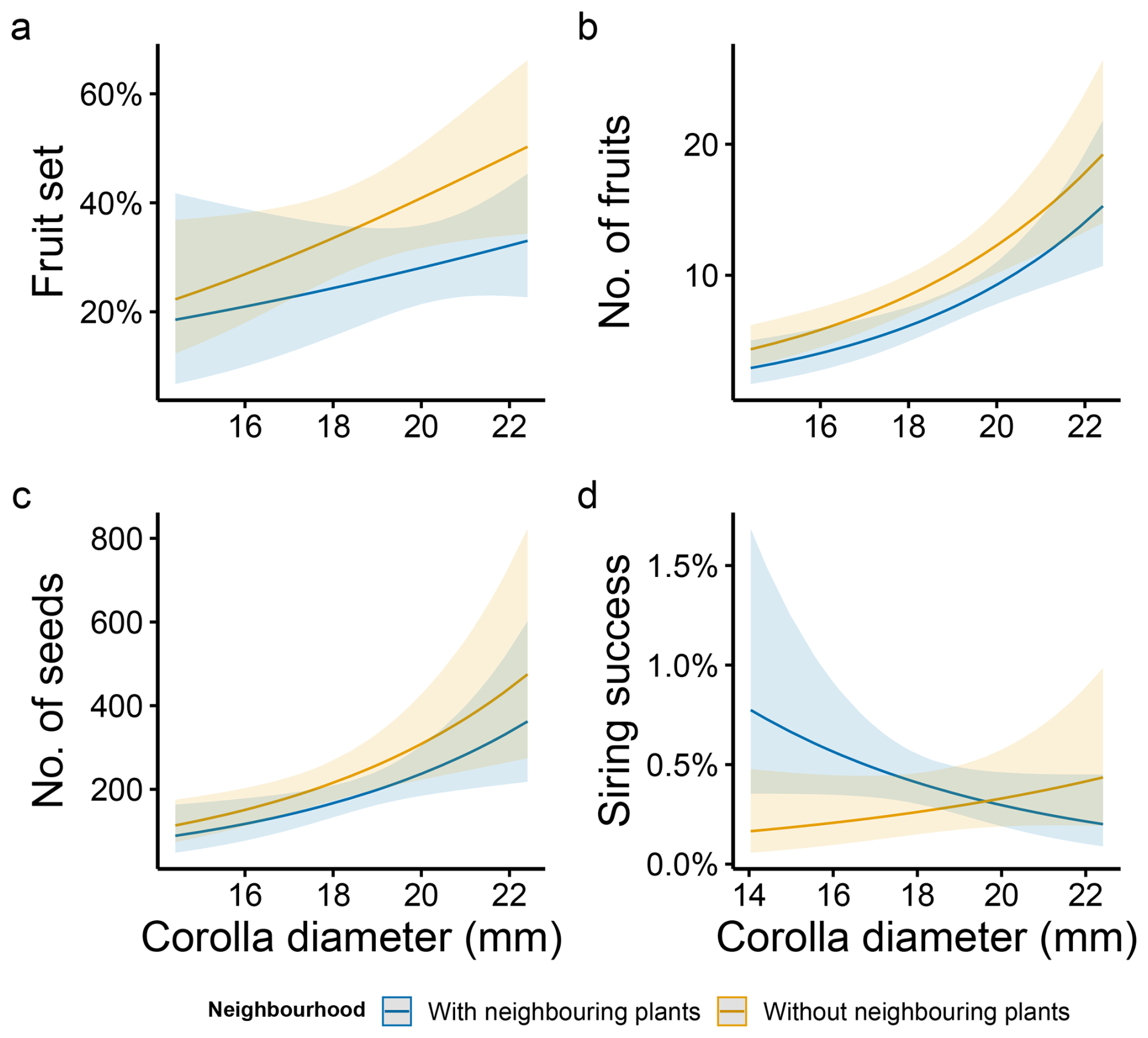
Figure 3Neighbourhood effect on the relationship between corolla size and female and male fitness components in M. moricandioides: (a) fruit set, (b) number of fruits, (c) number of seeds, and (d) siring success. Lines indicate mean values (±95 % CI) predicted by the model, and shaded areas indicate confidence intervals from the fitted model. Neighbourhood categories correspond to plants with/without conspecific neighbouring plants.
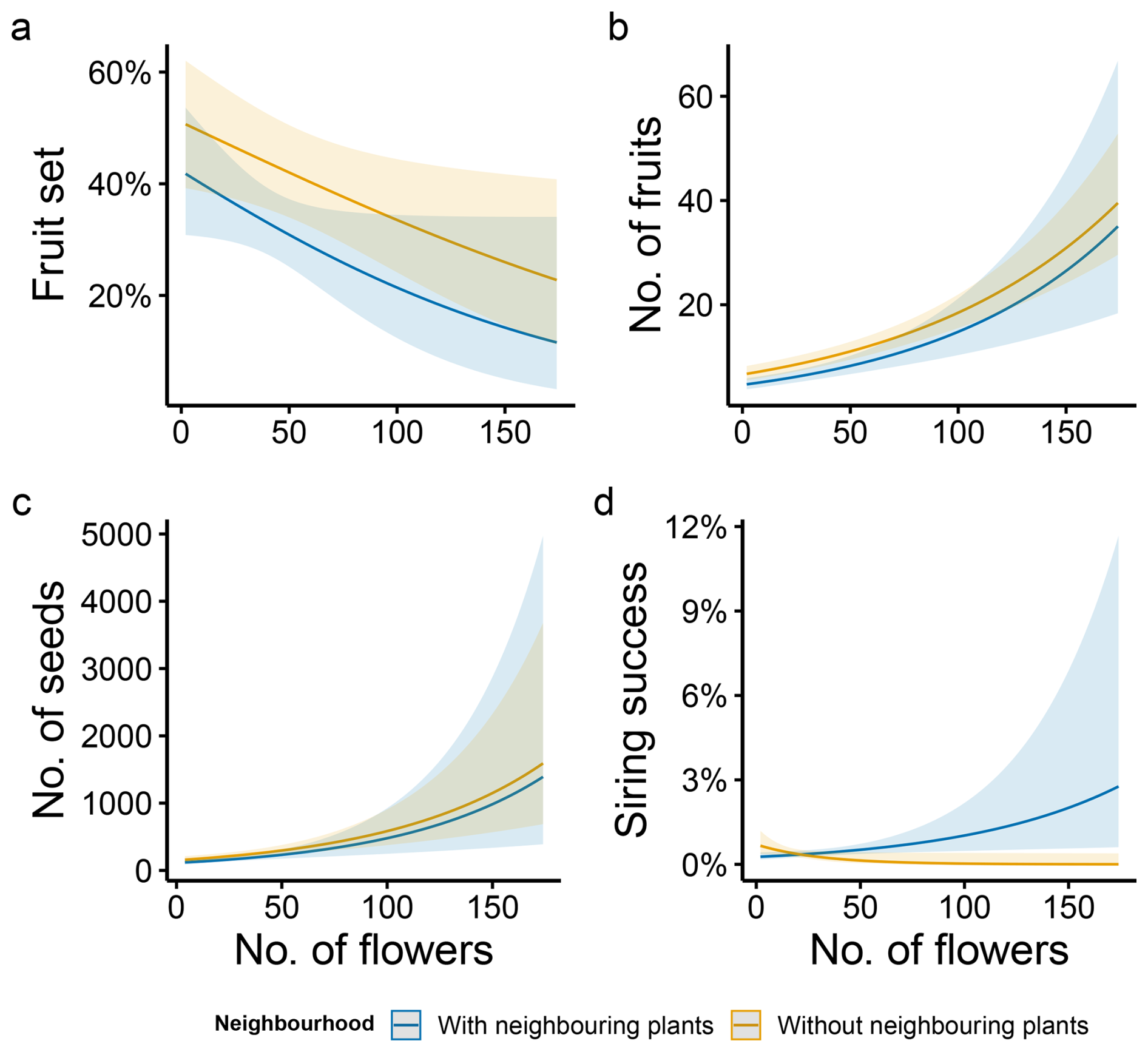
Figure 4Neighbourhood effect on the relationship between flower number and female and male fitness components in M. moricandioides: (a) fruit set, (b) number of fruits, (c) number of seeds, and (d) siring success for different values of number of flowers per plant. Lines indicate mean values (±95 % CI) predicted by the model, and shaded areas indicate confidence intervals from the fitted model. Neighbourhood categories correspond to plants with/without neighbouring conspecifics.
Siring success was otherwise significantly shaped by the number of flowers, and the interaction between both plant floral traits and the presence of neighbouring plants (Table 1, Figs. 3d and 4d). A contrasting pattern was found for flower number and corolla diameter, depending on whether plants grew alone or were surrounded by conspecifics. For plants growing without neighbouring individuals, corolla size was positively related to siring success, whereas for those growing in the presence of conspecifics, the relationship was negative (Fig. 3d). Conversely, the number of flowers was positively associated with siring success only among plants growing with conspecific neighbours, but for those growing without neighbouring plants, this relationship shifted to slightly negative (Fig. 4d).
3.2.2 Effect of the number of neighbours and its genetic relatedness on fitness
The number of neighbouring plants and the degree of genetic relatedness between them had different effects depending on the fitness component assessed. The number of conspecifics in the local neighbourhood significantly reduced fruit set but had no significant effect on the other female fitness components or their interactions (Table 2; Fig. 5a–c). In turn, the mean relatedness coefficient significantly reduced siring success (Table 2, Fig. 5d).
Table 2Effects of the number of neighbouring plants, the degree of genetic relatedness, and the interaction between them on female and male reproductive success as well as mean mating distances. Main effects were assessed using type III effects when the interaction was statistically significant, otherwise type II were used. The spatial coordinates were included as a random effect for the fruit set model. Variance and SD is shown for this random effect. Significant P values (<0.05) are highlighted in bold.
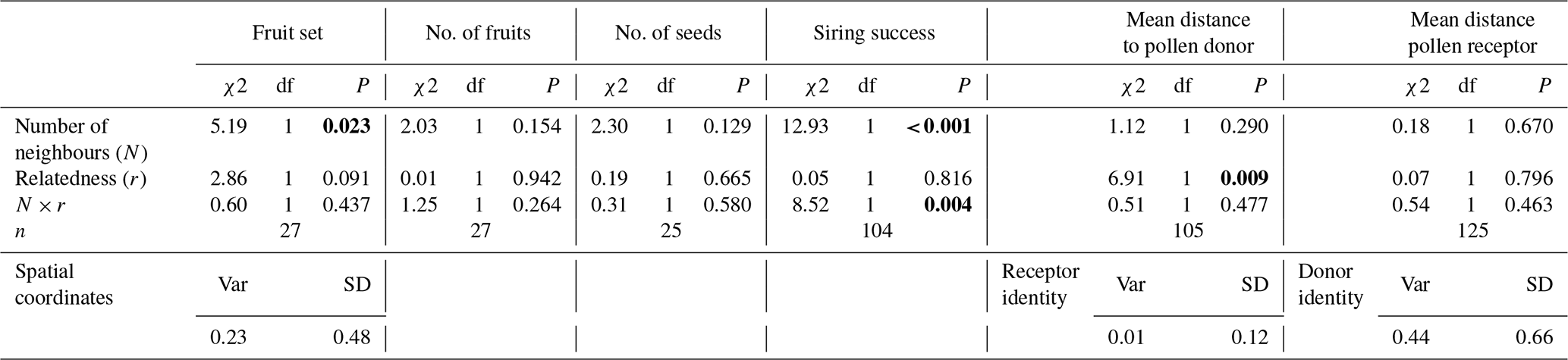
Figure 5Effects of the number of neighbouring plants and the degree of genetic relatedness between them on reproductive performance in M. moricandioides: (a) fruit set, (b) number of fruits, (c) number of seeds, and (d) siring success. Lines indicate mean values (±95 % CI) predicted by the model, and shaded areas indicate confidence intervals from the fitted model. Neighbourhood categories correspond to model predictions at the first quartile (Q1) and third quartile (Q3) values of relatedness coefficient distribution.
3.2.3 Neighbourhood effects on mating patterns
The mean distance between maternal plants and the assigned fathers within the study plot was 6.99 m ± 3.63 SD (Fig. 6a). The distribution of spatial distances between all assigned mating pairs did not differ significantly from the null distribution of pairwise spatial distances randomly sampled among all individuals in the plot (Fig. 6b). This result indicates that the distance between mating events occurred at the same frequency as the expected distribution of distances between individuals in this site.
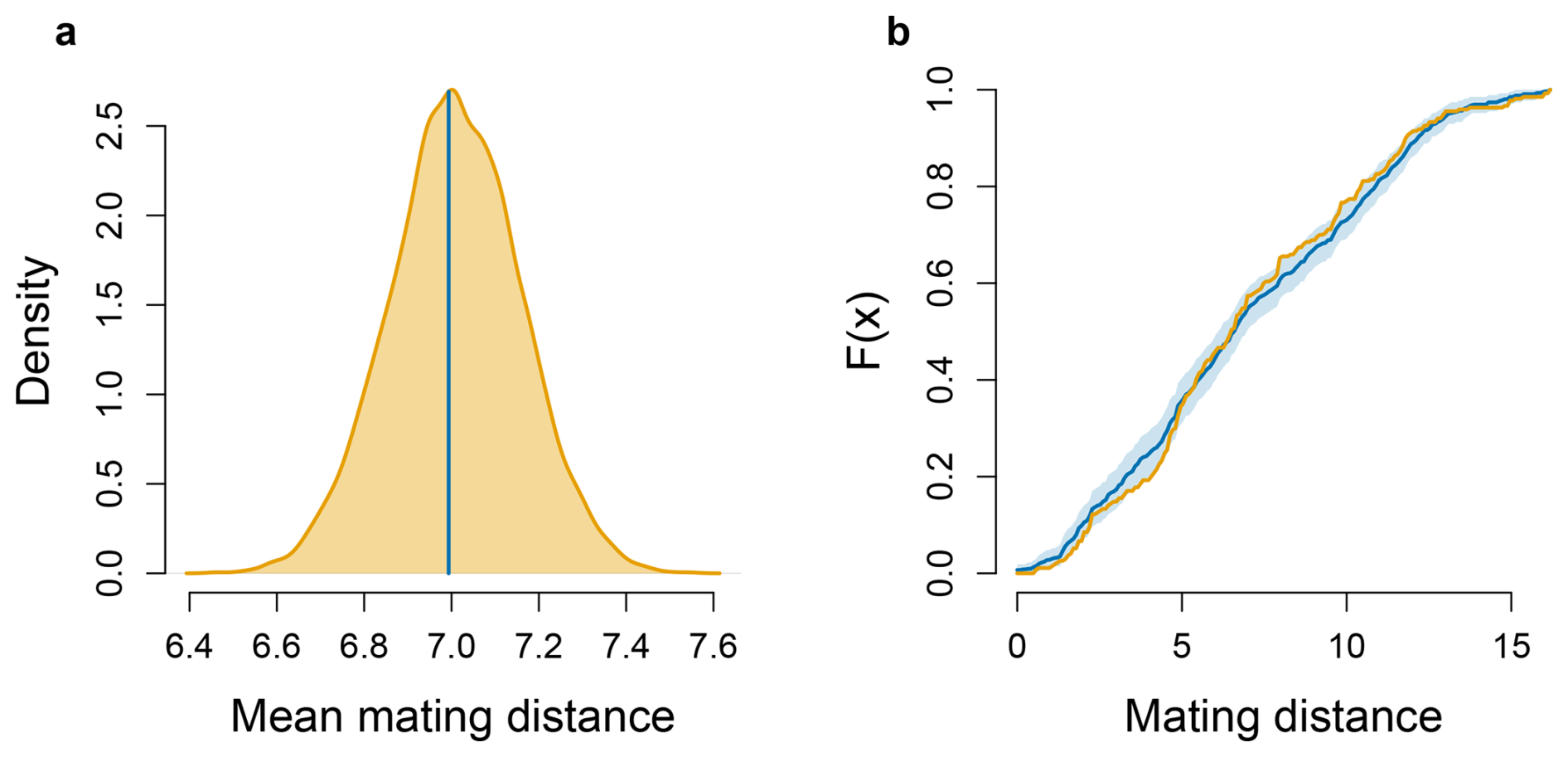
Figure 6Comparison between observed and null expectations of mating distances in M. moricandioides: (a) null 10 000 bootstrap distribution of the mean mating distance (yellow curve), with the observed mean indicated by the vertical blue line; (b) empirical cumulative distribution functions (ECDFs) of mating distances, with the yellow line representing observed mating pairs, and the blue line and shaded area indicating the null expectation (randomized pairs, mean ±95 % CI).
Floral traits and the presence of neighbouring conspecifics had no significant effect on the mean distance to the pollen donors (Table 1). However, for individuals with at least one conspecific neighbour within the local neighbourhood, the mean distance to the pollen donors increased significantly with the average pairwise relatedness coefficient of neighbouring individuals, showing no effect on the mean distance to the father (Table 2, Fig. 7).
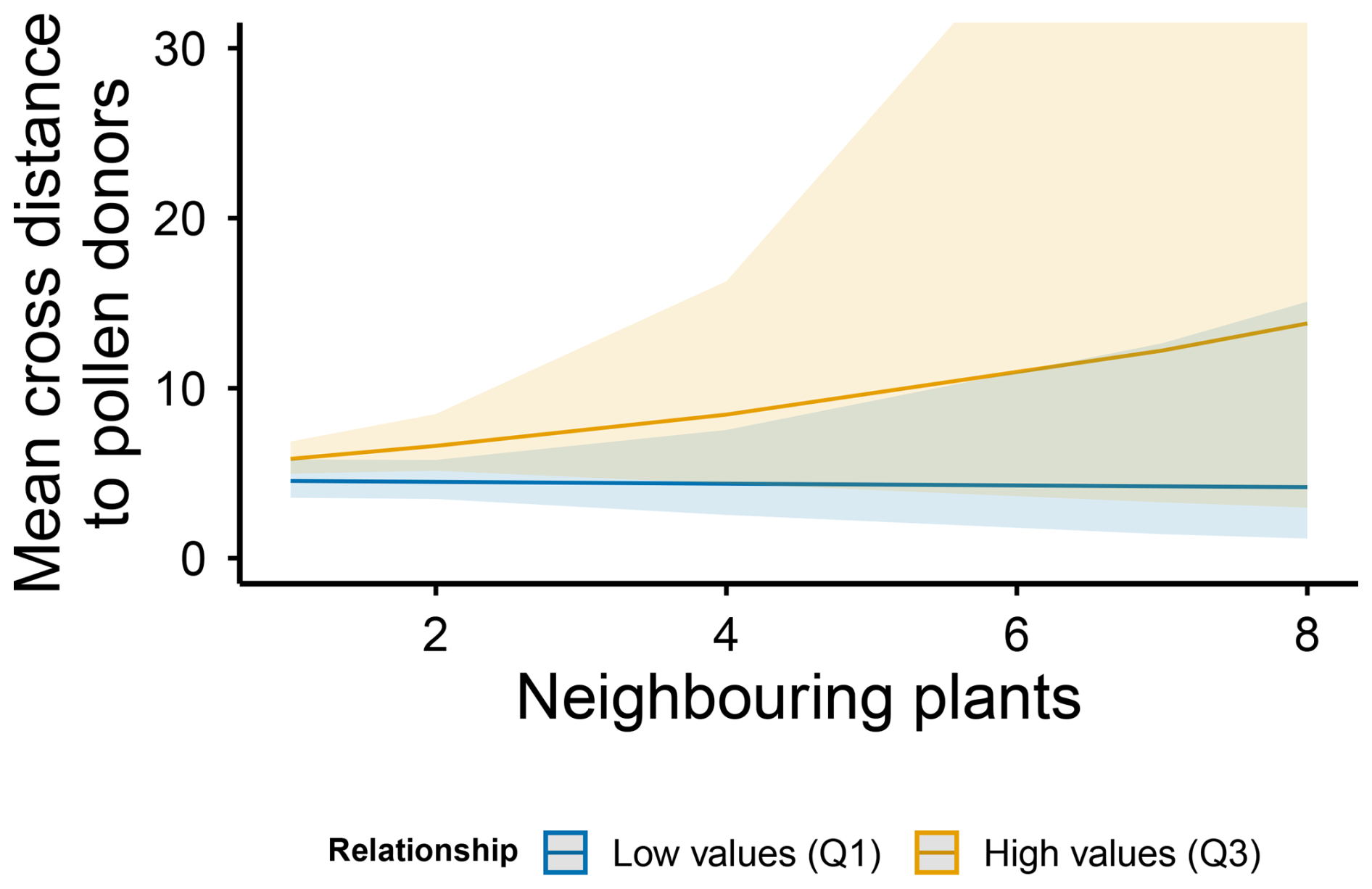
Figure 7Effects of neighbour number and genetic relatedness on mean cross distance to the pollen donors in M. moricandioides. Lines indicate mean values (±95 % CI) predicted by the model, and shaded areas indicate confidence intervals from the fitted model. Neighbourhood categories correspond to model predictions at the first quartile (Q1) and third quartile (Q3) values of relatedness coefficient distribution.
Our study supports the fact that the fine-scale spatial structure in a M. moricandioides population influenced reproductive success in complex and trait-dependent ways. We detected a spatial genetic structure consistent with isolation by distance, likely shaped by its limited seed dispersal, outcrossing the mating system and insect-mediated pollination. Such patterns are commonly observed in animal-pollinated species, where restricted pollen movement leads to local genetic clustering (Gamba and Muchhala, 2023; Vekemans and Hardy, 2004; Volis et al., 2016). In fact, the presence of conspecific neighbours affected both fitness components (female and male), while the direction and magnitude of these effects varied depending on floral display traits. Although neither the abundance nor the genetic relatedness of neighbouring plants significantly affected fruit or seed number, these neighbourhood attributes influenced fruit set and male fecundity, suggesting that genetic spatial aggregation may have important implications for specific components of plant fitness.
The presence of conspecific plants in the local neighbourhood had contrasting effects on different components of reproductive success. Female fitness components were consistently reduced in the presence of neighbours, regardless of floral traits, indicating that local neighbourhood effects may limit female reproductive output, possibly through competitive interference for resources (Mack and Harper, 1977; Silander and Pacala, 1985; Weiner, 1988; Miller and Schemske, 1990). Accordingly, plants with larger corollas or a greater number of flowers – traits potentially linked to higher resource status – showed increased female reproductive success, independently of the local neighbourhood context. In contrast, the effect of local neighbourhoods on male reproductive success depended on plant floral traits. The presence of neighbouring plants increased siring success in individuals with more flowers but not in those with larger corollas. Conversely, in solitary plants, siring success was higher in those with larger corollas but not in those producing more flowers. These findings contradict theoretical predictions regarding floral display traits under varying flower density (Cohen and Shmida, 1993), which propose that increased floral advertising through floral display should be more advantageous in low-density contexts where pollinator activity might be more limited. Meanwhile, in high-density patches, those traits enhancing pollinator choice, such as corolla size, may be favoured. However, several factors may alter these predictions. For instance, isolated plants may experience stronger negative effects of longer bouts associated with an increased flower number – a phenomenon known as the “attractiveness dilemma” (Klinkhamer and de Jong, 1993). Moreover, in self-incompatible species, increased intra-plant visitation by the same pollinator may lead to both pollen discounting, as pollen is deposited on stigmas of the same individual (Klinkhamer et al., 1994), and inhibition of the female function potentially reduces the effectiveness of subsequently arriving cross-pollen (Aizen and Harder, 2007; Snow et al., 1996). Under such low-density contexts, plants might enhance pollen export through advertising strategies other than increasing the number of simultaneously open flowers.
The effect of floral display traits on siring success when plants are surrounded by conspecifics will ultimately depend on the foraging behaviour of pollinators. Pollinators may leave a plant when the cost of revisiting flowers exceeds the cost of moving to another plant (Ohashi and Yahara, 2002), leading to a reduction in per-flower visitation rates under high flower density (e.g. Torices et al., 2026). Yet, plant aggregation may not affect all individuals equally, and group effects may depend on individual traits. For example, Ohashi and Yahara (2002) found that the number of flowers visited per plant increased less than proportionally with display size, and this increase was weaker at higher plant densities. Similarly, Grindeland et al. (2005) reported that the proportion of flowers visited decreased with display size, with a stronger decline in dense patches. Consequently, small displays received proportionally more visits in dense patches, while larger displays were favoured in sparse ones. Although our study lacked pollinator observations to directly assess these effects, siring success patterns suggest the opposite: plants with more flowers in dense neighbourhoods showed increased siring success. This may reflect a higher probability of attracting more insects to the group, rather than increased per-flower visitation rates. While our approach does not allow us to solve this puzzle, it underscores the importance of jointly considering individual traits, spatial aggregation, and neighbour characteristics when investigating the evolution of floral traits under pollinator-mediated selection.
Consistent with our second prediction, higher relatedness within the maternal neighbourhood was associated with greater mean mating distances and reduced siring success. In annual, entomophilous, and self-incompatible species such as M. moricandioides, a pronounced local genetic structure may constrain reproductive success due to compatibility limitations. The combination of spatial genetic structure and a self-incompatibility system likely reduces the success of matings among related neighbouring plants, thereby decreasing male siring success and potentially limiting female reproductive output. Increased mating distances may therefore reflect pollen movement beyond genetically similar neighbourhoods. Similar patterns have been reported in other annual and self-incompatible species, such as Raphanus sativus, where a higher relatedness degree led to reduced fitness through lower fruit set and seeds per fruit (Elam et al., 2007), and for Chamaecrista fasciculata where offspring fitness was lower when parents were neighbours than when they were more distant (Fenster, 1991). Moreover, these effects might be amplified by the negative consequences of biparental inbreeding depression on the offspring resulting from matings between related parents (Charlesworth and Charlesworth, 1987; Charlesworth and Willis, 2009; Jones and Comita, 2008).
Our results provide mixed support for the hypothesis that collective floral display among kin may enhance male reproductive success in M. moricandioides (Torices et al., 2018). This hypothesis proposes that the increased floral display previously observed in kin groups is a result of kin selection, where sharing pollinators among genetically related neighbours confers indirect fitness benefits through male reproductive success. In our study, we were unable to fully test this hypothesis, as we only evaluated the effects of individual traits on direct fitness components. Although we did not measure the traits of neighbouring plants, we did assess neighbourhood attributes such as presence, abundance, and genetic composition. Despite these limitations, we found that an increase in floral display – mediated by a higher number of flowers – when surrounded by other conspecifics did lead to increased male reproductive success, partially supporting the hypothesis. However, we found no evidence that higher genetic relatedness among neighbours enhanced male reproductive success; rather, it had the opposite effect. Interestingly, the positive influence of neighbourhood genetic structure was detected through female reproductive success: plants surrounded by more closely related neighbours tended to mate with pollen donors located at greater distances, potentially increasing the genetic diversity of their offspring. The mechanism underlying this pattern remains unclear. However, in self-incompatible species, the presence of closely related neighbours might act as a compatibility filter, reducing the likelihood of successful fertilization by locally available pollen. Under such conditions, compatible pollen arriving from more genetically distant individuals may be more likely to achieve fertilization, potentially explaining the increase in observed mating distances. Nevertheless, this result, together with the absence of negative effects of genetic structure on fruit or seed production, suggests that M. moricandioides may possess mechanisms to mitigate the potential female reproductive costs associated with high genetic relatedness in the local mating environment.
This study provides empirical evidence that fine-scale spatial and genetic structure within plant populations can shape reproductive outcomes in complex and trait-dependent ways. By characterizing the genetic landscape of a natural population of M. moricandioides, we identified a clear pattern of isolation by distance, consistent with passive seed dispersal and insect-mediated pollination. Our findings also revealed that the presence of conspecific neighbours modulates the relationship between floral display traits and reproductive success. While solitary plants consistently exhibited higher female fitness, male fitness was influenced by both neighbourhood context and individual-level flower size and number. These results underscore the importance of considering spatial context when evaluating the selective pressures acting on reproductive traits. Finally, we show that neighbourhood density and genetic relatedness can affect specific components of reproductive success. Notably, plants in genetically related neighbourhoods showed lower siring success, yet tended to mate with more distant pollen donors, indicating that local genetic structure may influence mating patterns and potentially enhance offspring genetic diversity. Although the mechanisms underlying these patterns remain unclear, our results contribute to a growing body of research highlighting the ecological relevance of spatial and genetic structure in shaping plant reproductive strategies.
All raw data can be provided by the corresponding authors upon request.
The supplement related to this article is available online at https://doi.org/10.5194/we-26-71-2026-supplement.
CF, RT, VQA, CP, IA, LGQ, LGB, and AJMP planned the campaign. CF, RT, VQA, CP, IA, LGQ and LGB performed the measurements. CF and RT analysed the data. CF and RT wrote the paper draft. CF, RT, CP, IA, VQA, LGQ, LGB, and AJMP reviewed and edited the article.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We thank Marcelino de la Cruz for assistance with experimental design and data analysis; and Kai-Hsiu Chen, Ehouarn Le Faou, John Pannell, and Alejandro Llanos-Garrido for their help with genomic data analyses. We also thank Adrian Brennan, Mason Kulbaba, and one anonymous reviewer for their comments on the manuscript. The authors acknowledge the assistance of OpenAI's ChatGPT (GPT-5, 2025) for language editing, formatting guidance, and technical support during the preparation of figures and statistical descriptions. All scientific interpretations, analyses, and conclusions are the authors' own.
This work was funded by the Spanish State Research Agency (10.13039/501100011033) through the 2020 Call of the I+D+I Projects for Knowledge Generation (grant no. PID2020-113102GA-I00) and an FPI predoctoral contract (grant no. PRE2021-099387), and partially by RARABUN 2022/00156/001 (Comunidad de Madrid).
This paper was edited by Adrian Brennan and reviewed by Mason Kulbaba and one anonymous referee.
Aizen, M. A. and Harder, L. D.: Expanding the Limits of the Pollen-Limitation Concept: Effects of Pollen Quantity and Quality, Ecology, 88, 271–281, https://doi.org/10.1890/06-1017, 2007.
Bates, D., Mächler, M., Bolker, B., and Walker, S.: Fitting linear mixed-effects models using lme4, J. Stat. Softw., 67, 1–48, https://doi.org/10.18637/jss.v067.i01, 2015.
Biernaskie, J. M., Walker, S. C., and Gegear, R. J.: Bumblebees Learn to Forage like Bayesians, Am. Nat., 174, 413–423, https://doi.org/10.1086/603629, 2009.
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., Skaug, H. J., Maechler, M., and Bolker, B. M.: glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling, R J., 9, 378–400, https://doi.org/10.32614/RJ-2017-066, 2017.
Charlesworth, D. and Charlesworth, B.: Inbreeding depression and its evolutionary consequences, Annu. Rev. Ecol. Evol. S., 18, 237–268, https://doi.org/10.1146/annurev.es.18.110187.001321, 1987.
Charlesworth, D. and Willis, J. H.: The genetics of inbreeding depression, Nat. Rev. Genet., 10, 783–796, https://doi.org/10.1038/nrg2664, 2009.
Cohen, D. and Shmida, A.: The Evolution of Flower Display and Reward, Evol. Biol., 27, 197–243, https://doi.org/10.1007/978-1-4615-2878-4_6, 1993.
Danecek, P., Auton, A., Abecasis, G., Albers, C. A., Banks, E., DePristo, M. A., Handsaker, R. E., Lunter, G., Marth, G. T., Sherry, S. T., McVean, G., Durbin, R., and 1000 Genomes Project Analysis Group: The variant call format and VCFtools, Bioinformatics, 27, 2156–2158, https://doi.org/10.1093/bioinformatics/btr330, 2011.
Danecek, P., Bonfield, J. K., Liddle, J., Marshall, J., Ohan, V., Pollard, M. O., Whitwham, A., Keane, T., McCarthy, S. A., Davies, R. M., and Li, H.: Twelve years of SAMtools and BCFtools, GigaScience, 10, giab008, https://doi.org/10.1093/gigascience/giab008, 2021.
Dauber, J., Biesmeijer, J. C., Gabriel, D., Kunin, W. E., Lamborn, E., Meyer, B., Nielsen, A., Potts, S. G., Roberts, S. P. M., Sõber, V., Settele, J., Steffan-Dewenter, I., Stout, J. C., Teder, T., Tscheulin, T., Vivarelli, D., and Petanidou, T.: Effects of patch size and density on flower visitation and seed set of wild plants: a pan-European approach, J. Ecol., 98, 188–196, https://doi.org/10.1111/j.1365-2745.2009.01590.x, 2010.
Doyle, J. J. and Doyle, J. L.: A rapid DNA isolation procedure for small quantities of fresh leaf tissue, Phytochemical Bulletin, 19, 11–15, 1987.
Dutech, C., Sork, V. L., Irwin, A. J., Smouse, P. E., and Davis, F. W.: Gene flow and fine-scale genetic structure in a wind-pollinated tree species, Quercus lobata (Fagaceaee), Am. J. Bot., 92, 252–261, https://doi.org/10.3732/ajb.92.2.252, 2005.
Elam, D. R., Ridley, C. E., Goodell, K., and Ellstrand, N. C.: Population size and relatedness affect fitness of a self-incompatible invasive plant, P. Natl. Acad. Sci. USA, 104, 549–552, https://doi.org/10.1073/pnas.0607306104, 2007.
Epperson, B. K.: Spatial structure of genetic variation within populations of forest trees, New Forest, 6, 257–278, https://doi.org/10.1007/BF00120648, 1992.
Escudero, A., Iriondo, J. M., and Torres, M. E.: Spatial analysis of genetic diversity as a tool for plant conservation, Biol. Conserv., 113, 351–365, https://doi.org/10.1016/S0006-3207(03)00122-8, 2003.
Fenster, C. B.: Gene Flow in Chamaecrista Fasciculata (leguminosae) I. Gene Dispersal, Evolution, 45, 398–409, https://doi.org/10.1111/j.1558-5646.1991.tb04413.x, 1991.
Gamba, D. and Muchhala, N.: Pollinator type strongly impacts gene flow within and among plant populations for six Neotropical species, Ecology, 104, e3845, https://doi.org/10.1002/ecy.3845, 2023.
Ghazoul, J.: Pollen and seed dispersal among dispersed plants, Biol. Rev., 80, 413–443, https://doi.org/10.1017/S1464793105006731, 2005.
Gómez, J. M.: Predispersal reproductive ecology of an arid land crucifer, Moricandia moricandioides: effect of mammal herbivory on seed production, J. Arid Environ., 33, 425–437, https://doi.org/10.1006/jare.1996.0078, 1996.
Gómez, J. M., Torices, R., Lorite, J., Klingenberg, C. P., and Perfectti, F.: The role of pollinators in the evolution of corolla shape variation, disparity and integration in a highly diversified plant family with a conserved floral bauplan, Ann. Bot., 117, 889–904, https://doi.org/10.1093/aob/mcv194, 2016.
Grindeland, J. M., Sletvold, N., and Ims, R. A.: Effects of Floral Display Size and Plant Density on Pollinator Visitation Rate in a Natural Population of Digitalis purpurea, Funct. Ecol., 19, 383–390, 2005.
Hardy, O. J. and Vekemans, X.: spagedi: a versatile computer program to analyse spatial genetic structure at the individual or population levels, Mol. Ecol. Notes, 2, 618–620, https://doi.org/10.1046/j.1471-8286.2002.00305.x, 2002.
Hardy, O. J., Maggia, L., Bandou, E., Breyne, P., Caron, H., Chevallier, M.-H., Doligez, A., Dutech, C., Kremer, A., Latouche-Hallé, C., Troispoux, V., Veron, V., and Degen, B.: Fine-scale genetic structure and gene dispersal inferences in 10 Neotropical tree species, Mol. Ecol., 15, 559–571, https://doi.org/10.1111/j.1365-294X.2005.02785.x, 2006.
Hartig, F.: DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models, R package version 0.4.6, https://CRAN.R-project.org/package=DHARMa (last access: 19 May 2026), 2022.
Hegland, S. J.: Floral neighbourhood effects on pollination success in red clover are scale-dependent, Funct. Ecol., 28, 561–568, https://doi.org/10.1111/1365-2435.12223, 2014.
Huisman, J.: Pedigree reconstruction from SNP data: parentage assignment, sibship clustering and beyond, Mol. Ecol. Resour., 17, 1009–1024, https://doi.org/10.1111/1755-0998.12665, 2017.
Hurtado, M., Godoy, O., and Bartomeus, I.: Plant spatial aggregation modulates the interplay between plant competition and pollinator attraction with contrasting outcomes of plant fitness, Web Ecol., 23, 51–69, https://doi.org/10.5194/we-23-51-2023, 2023.
Jones, F. A. and Comita, L. S.: Neighbourhood density and genetic relatedness interact to determine fruit set and abortion rates in a continuous tropical tree population, P. Roy. Soc. B-Biol. Sci., 275, 2759–2767, https://doi.org/10.1098/rspb.2008.0894, 2008.
Jones, F. A. and Hubbell, S. P.: Demographic spatial genetic structure of the Neotropical tree, Jacaranda copaia, Mol. Ecol., 15, 3205–3217, https://doi.org/10.1111/j.1365-294X.2006.03023.x, 2006.
Klinkhamer, P. G. L. and de Jong, T. J.: Attractiveness to pollinators: a plant's dilemma, Oikos, 66, 180–184, 1993.
Klinkhamer, P. G. L., de Jong, T. J., and Metz, J. A. J.: Why plants can be too attractive – A discussion of measures to estimate male fitness, J. Ecol., 82, 191–194, 1994.
Klinkhamer, P. G. L., De Jong, T. J., and Linnebank, L. A.: Small-scale spatial patterns determine ecological relationships: an experimental example using nectar production rates, Ecol. Lett., 4, 559–567, https://doi.org/10.1046/j.1461-0248.2001.00267.x, 2001.
Knight, T. M.: Floral density, pollen limitation, and reproductive success in Trillium grandiflorum, Oecologia, 137, 557–563, https://doi.org/10.1007/s00442-003-1371-8, 2003.
Larridon, I., Villaverde, T., Zuntini, A. R., Pokorny, L., Brewer, G. E., Epitawalage, N., Fairlie, I., Hahn, M., Kim, J., Maguilla, E., Maurin, O., Xanthos, M., Hipp, A. L., Forest, F., and Baker, W. J.: Tackling Rapid Radiations With Targeted Sequencing, Front. Plant Sci., 10, https://doi.org/10.3389/fpls.2019.01655, 2020.
Laverty, T. M.: Plant interactions for pollinator visits: a test of the magnet species effect, Oecologia, 89, 502–508, https://doi.org/10.1007/BF00317156, 1992.
Lenth, R. V.: emmeans: Estimated Marginal Means, aka Least-Squares Means, R package version 1.10.4, https://CRAN.R-project.org/package=emmeans (last access: 19 May 2026), 2024.
Li, H.: Aligning sequence reads, clone sequences and assembly contigs with BWA-MEM, arXiv [preprint], https://doi.org/10.48550/arXiv.1303.3997, 26 May 2013.
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., Marth, G., Abecasis, G., and Durbin, R.: The Sequence Alignment/Map format and SAMtools, Bioinformatics, 25, 2078–2079, https://doi.org/10.1093/bioinformatics/btp352, 2009.
Loiselle, B. A., Sork, V. L., Nason, J., and Graham, C.: Spatial Genetic Structure of a Tropical Understory Shrub, Psychotria officinalis (Rubiaceae), Am. J. Bot., 82, 1420–1425, https://doi.org/10.2307/2445869, 1995.
Loveless, M. D. and Hamrick, J. L.: Ecological Determinants of Genetic Structure in Plant Populations, Annu. Rev. Ecol. Syst., 15, 65–95, https://doi.org/10.1146/annurev.es.15.110184.000433, 1984.
Lüdecke, D.: ggeffects: Create Tidy Data Frames of Marginal Effects for ggplot from Model Outputs, R package version 1.7.2, https://CRAN.R-project.org/package=ggeffects (last access: 19 May 2026), 2023.
Mack, R. N. and Harper, J. L.: Interference in Dune Annuals: Spatial Pattern and Neighbourhood Effects, J. Ecol., 65, 345, https://doi.org/10.2307/2259487, 1977.
Martén-Rodríguez, S., Quesada, M., and Momen, B.: Effects of Local Plant Density and Incomplete Dichogamy on the Reproductive Success of the Rare Neotropical Palm Geonoma epetiolata, Biotropica, 44, 680–688, https://doi.org/10.1111/j.1744-7429.2012.00860.x, 2012.
Miller, T. E. and Schemske, D. W.: An Experimental Study of Competitive Performance in Brassica rapa (Cruciferae), Am. J. Bot., 77, 993, https://doi.org/10.2307/2444570, 1990.
Moeller, D. A.: Facilitative Interactions Among Plants Via Shared Pollinators, Ecology, 85, 3289–3301, https://doi.org/10.1890/03-0810, 2004.
Nathan, R. and Muller-Landau, H. C.: Spatial patterns of seed dispersal, their determinants and consequences for recruitment, Trends Ecol. Evol., 15, 278–285, https://doi.org/10.1016/S0169-5347(00)01874-7, 2000.
Nogueira, F. C. B., Filho, S. M., Baldoni, R. N., and Sampaio e Silva, T. A.: Is the Seed Dispersal Related to Spatial Pattern of Individuals in Populations? The Case of Dalbergia cearensis, American Journal of Plant Sciences, 5, 2997–3004, https://doi.org/10.4236/ajps.2014.520316, 2014.
Ohashi, K. and Yahara, T.: Visit larger displays but probe proportionally fewer flowers: counterintuitive behaviour of nectar-collecting bumble bees achieves an ideal free distribution, Funct. Ecol., 16, 492–503, https://doi.org/10.1046/j.1365-2435.2002.00644.x, 2002.
Paradis, E. and Schliep, K.: ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R, Bioinformatics, 35, 526–528, https://doi.org/10.1093/bioinformatics/bty633, 2019.
Peakall, R. and Beattie, A. J.: Does ant dispersal of seeds in Sclerolaena diacantha (Chenopodiaceae) generate local spatial genetic structure?, Heredity, 75, 351–361, https://doi.org/10.1038/hdy.1995.146, 1995.
Purcell, S., Neale, B., Todd-Brown, K., Thomas, L., Ferreira, M. A. R., Bender, D., Maller, J., Sklar, P., de Bakker, P. I. W., Daly, M. J., and Sham, P. C.: PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses, Am. J. Hum. Genet., 81, 559–575, https://doi.org/10.1086/519795, 2007.
Queller, D. C. and Goodnight, K. F.: Estimating relatedness using genetic markers, Evolution, 43, 258–275, https://doi.org/10.1111/j.1558-5646.1989.tb04226.x, 1989.
R Core Team: R: The R Project for Statistical Computing, https://www.r-project.org/ (last access: 19 May 2026), 2026.
Ruttan, A., Lortie, C. J., and Haas, S. M.: Shrubs as magnets for pollination: A test of facilitation and reciprocity in a shrub-annual facilitation system, Current Research in Insect Science, 1, 100008, https://doi.org/10.1016/j.cris.2021.100008, 2021.
Schupp, E. W., Jordano, P., and Gómez, J. M.: Seed dispersal effectiveness revisited: a conceptual review, New Phytol., 188, 333–353, https://doi.org/10.1111/j.1469-8137.2010.03402.x, 2010.
Silander, J. A. and Pacala, S. W.: Neighborhood predictors of plant performance, Oecologia, 66, 256–263, https://doi.org/10.1007/bf00379863, 1985.
Snow, A. A., Spira, T. P., Simpson, R., and Klips, R. A.: The Ecology of Geitonogamous Pollination, Floral Biology, https://doi.org/10.1007/978-1-4613-1165-2_7, 1996.
Sobrino Vesperinas, E.: Moricandia DC., in: Flora Iberica, edited by: Castroviejo, S., CSIC, Madrid, 337–344, ISBN 84-00-07385-1, 1993.
Stacy, E. A., Hamrick, J. L., Nason, J. D., Hubbell, S. P., Foster, R. B., and Condit, R.: Pollen Dispersal in Low-Density Populations of Three Neotropical Tree Species, Am. Nat., 148, 275–298, 1996.
Sun, S., Broom, M., Johanis, M., and Rychtář, J.: A mathematical model of kin selection in floral displays, J. Theor. Biol., 509, 110470, https://doi.org/10.1016/j.jtbi.2020.110470, 2021.
Thomson, J. D.: Spatial and Temporal Components of Resource Assessment by Flower-Feeding Insects, J. Anim. Ecol., 50, 49–59, https://doi.org/10.2307/4030, 1981.
Torices, R., Gómez, J. M., and Pannell, J. R.: Kin discrimination allows plants to modify investment towards pollinator attraction, Nat. Commun., 9, 2018, https://doi.org/10.1038/s41467-018-04378-3, 2018.
Torices, R., González-Mejías, A., and Gómez, J. M.: Influence of group size and floral display on pollinator behaviour in Moricandia arvensis, J. Poll. Ecol., https://doi.org/10.26786/1920-7603(2026)882, 2026.
Underwood, N., Hambäck, P. A., and Inouye, B. D.: Pollinators, Herbivores, and Plant Neighborhood Effects, Q. Rev. Biol., 95, 37–57, https://doi.org/10.1086/707863, 2020.
Vekemans, X. and Hardy, O. J.: New insights from fine-scale spatial genetic structure analyses in plant populations, Mol. Ecol., 13, 921–935, https://doi.org/10.1046/j.1365-294X.2004.02076.x, 2004.
Volis, S., Ormanbekova, D., and Shulgina, I.: Fine-scale spatial genetic structure in predominantly selfing plants with limited seed dispersal: A rule or exception?, Plant Divers., 38, 59–64, https://doi.org/10.1016/j.pld.2016.03.001, 2016.
Weiner, J.: The influence of competition on plant reproduction, in: Plant Reproductive Ecology. Patterns and strategies, edited by: Lovett Doust, J. and Lovett-Doust, L., Oxford University Press, New York, 203–227, ISBN 0-19-506394-5, 1988.
Wickham, H.: ggplot2: Elegant Graphics for Data Analysis, Springer-Verlag, New York, https://doi.org/10.1007/978-3-319-24277-4, 2016.