the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 11 Jun 2026
| 11 Jun 2026
Blueberry floral morphology influences pollinator visitation and behavioural patterns
Ana Margarida Branco
Bruna Chaves
Hugo Gaspar
João Loureiro
Sílvia Castro
The blueberry is a globally important fruit crop with growing demand that depends on insect pollinators for market-quality fruits. However, pollination is far from a simple process and is affected by several interacting factors. Among these, flower morphology plays a key role in determining which visitors are pollinators, the visitation rates and the occurrence of nectar robbing. In this study, we seek to identify the main pollinators for several blueberry cultivars, and relate insect floral visitors and visitation rates to flower morphology. To this end, we conducted a study in an experimental field in the Centro region of Portugal, with 19 blueberry cultivars growing under the same conditions. Flower visitors were observed, and their identity and the number of flowers visited were recorded in 5 min periods distributed throughout the day. Flower morphological traits related to pollination were measured using 6–10 flowers per individual bush. Analysis of flower morphological traits reveals variation among cultivars. Data also indicate that flower morphology influences the behaviour of insect visitors. We found three main visitors: Apis mellifera, which tended to visit cultivars with smaller flowers; Anthophora plumipes, which tended to visit cultivars with larger flowers; and Bombus terrestris, which visited cultivars with intermediate-sized flowers. Overall, our findings indicate that cultivar flower morphology and pollinator proboscis size are important in blueberry pollination, and corolla size variation should be considered in the selection of cultivars for field design. Selecting cultivars that best suit local pollination services and pollination management strategy (managed pollinators or a natural pollinator community) is key to improving production.
- Article
(3105 KB) - Full-text XML
-
Supplement
(270 KB) - BibTeX
- EndNote




Over 75 % of crops rely partially or completely on pollination by animals for quality fruit and seed production, with pollinators representing USD195–387 billion per annum of the global food economy (Porto et al., 2020). Yet, pollination is far from being a simple process and is affected by several interacting factors (Castro et al., 2021; Gérard et al., 2020; Kuppler and Kotowska, 2021; Potts et al., 2010). The abundance and diversity of pollinator communities determine the levels of pollination services, while differences in traits and rewards among crop cultivars influence pollinators' behaviour based on their preferences and morphological traits (Arrington and DeVetter, 2018; Courcelles et al., 2013; Symington and Glover, 2024; Woodcock et al., 2019). Given the importance of successful pollination for crop yields and quality, understanding plant–pollinator interactions in crop systems and improving management practices are essential in optimising crop yields.
Blueberry (Vaccinium spp.) is a globally important fruit crop that encompasses multiple species, including northern highbush and southern highbush (V. corymbosum), and rabbiteye (complex hybrids including V. corymbosum and V. virgatum; Song and Hancock, 2011). Crop production depends on insect pollinators to produce market-quality fruits. If pollination is adequate and when resources are not limited, the highbush blueberry can potentially reach a fruit set of 100 % (Dogterom et al., 2000; Ehlenfeldt, 2001; Kumarihami et al., 2021). Although there are varying levels of self-fertility, dependent on species and cultivar, blueberries are primarily outcrossing, showing larger fruit size and earlier fruit ripening when cross-pollinated (Cortés-Rivas et al., 2023b; Dogterom et al., 2000; Song and Hancock, 2011; Taber and Olmstead, 2016). All these characteristics make insect pollination essential for blueberries. However, while there are several studies addressing blueberry cultivar selection, production and management, the investment in research in pollination management is much lower despite its importance in supporting evidence-based decision-making for sustainable production (DeVetter et al., 2022; Edger et al., 2022).
Pollination is affected by several interacting factors, including the abundance and richness of pollinators, flower morphology, floral attractants and rewards, and climatic conditions (Castro et al., 2021; Gérard et al., 2020; Kuppler and Kotowska, 2021; Potts et al., 2010). Considerable attention has been given to understanding the factors influencing pollinator abundance and diversity in blueberry fields (e.g. Cane and Payne, 1993; Eeraerts et al., 2023; Isaacs and Kirk, 2010; Nicholson et al., 2017). Some attention has also been given to pollinator efficiency (e.g., Cortés-Rivas et al., 2023b; Kendall et al., 2020, 2022; Payne et al., 1989) with stingless bees (tribe Meliponini – absent from Europe; Kendall et al., 2022) and bees capable of buzz pollination, such as bumblebees (Bombus spp., De Luca and Vallejo-Marín, 2013; Cortés-Rivas et al., 2023a) and southern blueberry bees (Habropoda laboriosa – absent from Europe; Payne et al., 1989), which are more efficient visitors due to their ability to buzz and/or faster flower handling time (Cane and Payne, 1988; Cortés-Rivas et al., 2023a). In contrast, the honey bee (Apis mellifera Linnaeus, 1758) requires multiple visits for complete pollination (Eeraerts et al., 2023). However, far less attention has been given to factors that attract and influence pollinator behaviour and, consequently, pollination success, such as variations in flower morphology (DeVetter et al., 2022), with only a few studies addressing the effect of floral morphology on insect visitors (e.g. Arrington and DeVetter, 2018; Courcelles et al., 2013; Sampson et al., 2013). Blueberry flowers have morphological characteristics that prevent the occurrence of autonomous self-pollination and, therefore, require insect pollination. Flowers are upside down and tubular. The anthers are poricidal, requiring buzz pollination for adequate pollen release (Vander Kloet, 1988). The stamens are shorter than the style, and the distance between the anthers and the stigma varies by cultivar, as does the stigmatic protrusion from the corolla (Courcelles et al., 2013; Vander Kloet, 1988). This phenotypic variation in flower morphology has been shown to affect insect behaviour and influence pollination success (Arrington and DeVetter, 2018; Courcelles et al., 2013; Sampson et al., 2013). The shape of blueberry flowers, combined with the positioning of the nectaries between the base of the style and the anther filaments, makes it difficult for insects to access the nectar and constrains pollination to specific insect species (Irwin et al., 2010; Stang et al., 2009).
Some studies have addressed the variation in flower size among blueberry cultivars (e.g. Arrington and DeVetter, 2018; Courcelles et al., 2013; Eck and Mainland, 1971; Sampson et al., 2013) but few have looked at the relationship between flower morphology and insect visits (e.g. Courcelles et al., 2013; Sampson et al., 2013). Courcelles et al. (2013) compared four northern highbush cultivars – Bluecrop, Draper, Duke and Liberty – and found that the diameter of the corolla opening was the main driver of honeybee visitation rates, with higher values recorded in cultivars with wider corolla openings. However, to our knowledge, no study has addressed the relationship between floral morphology and insect visitation across multiple cultivars of northern, southern and rabbiteye blueberry exposed to the same insect assemblage.
This study aimed to explore the impact of blueberry floral traits on pollinator visitation and behaviour. Because both floral and insect-pollinator traits vary, longer corollas and smaller apertures are expected to constrain floral visitors' access to nectar rewards. We hypothesise that cultivars with longer corollas and smaller apertures will impose greater constraints on insect pollinators, thereby influencing visitation rates. Shorter corollas are expected to be visited by a wider assemblage of floral visitors, while longer corollas are expected to be visited by insects with longer proboscises and be subject to higher rates of nectar robbing (Fig. 1). To test this, we measured the corollas of 19 blueberry cultivars and identified the flower visitors, their behaviour and visitation rates.
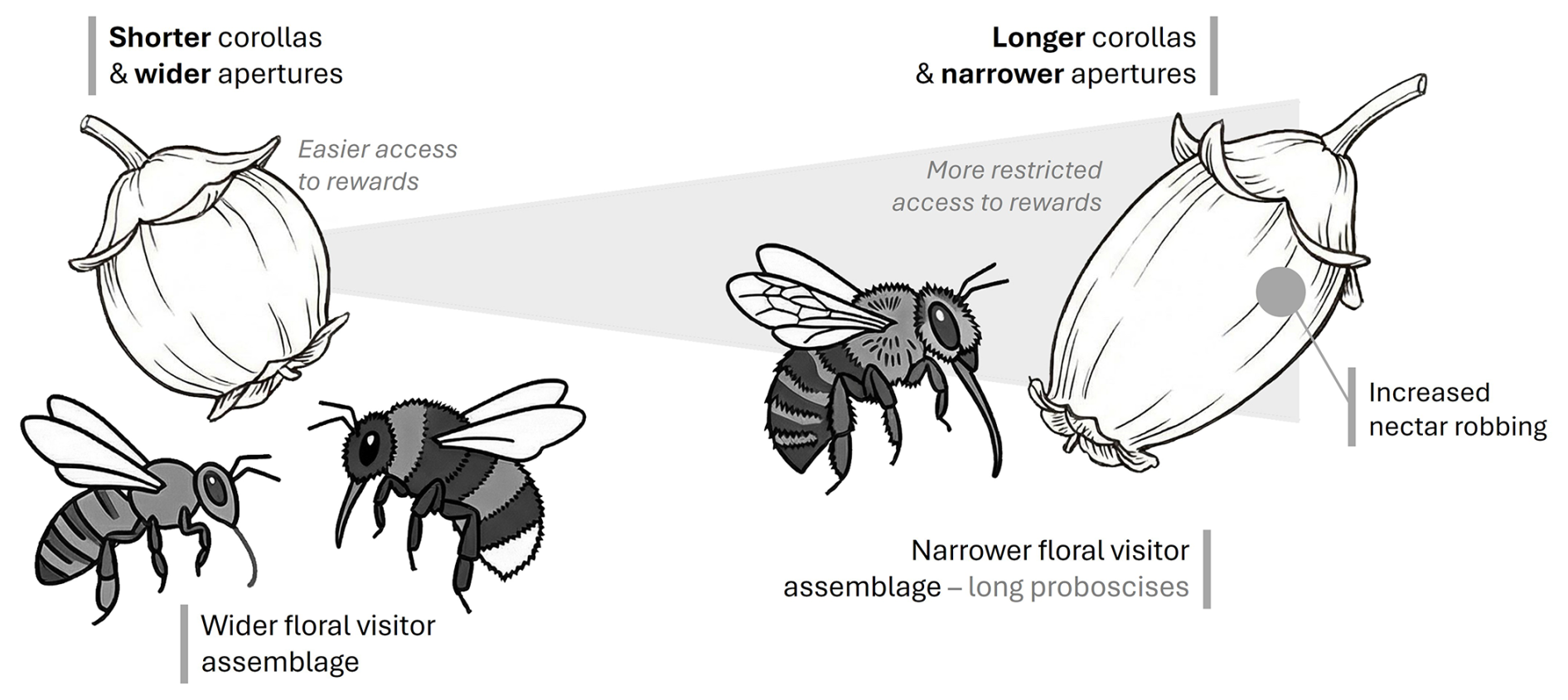
Figure 1Conceptual schematic representation of the hypothesis for plant–pollinator interactions linked to corolla morphology. We hypothesise that cultivars with shorter corollas will be visited by a wider assemblage of floral visitors due to easier accessibility to rewards (left side), while increasing corolla length (represented by the shaded area) will narrow the assemblage of potential legitimate visitors, so that cultivars with longer corollas are expected to be visited by insects with longer proboscises and be subject to higher rates of nectar robbing (right side). Original drawing by S. Castro, digitalised and further processed using AI-assisted tools, and finalised by the authors.
2.1 Study site
The study was carried out in a blueberry experimental field located at the Agrarian Field Station of Viseu, Portugal (40°39′52.4′′ N, 7°54′12.6′′ W). Nineteen blueberry cultivars were evaluated, including 10 northern highbush (“Aurora”, “Bluecrop”, “Bluegold”, “Chandler”, “Draper”, “Duke”, “Elliot”, “Huron”, “Legacy”, “Liberty”), eight southern highbush cultivars (“Biloxi”, “Camelia”, “Misty”, “O'Neil”, “Rebel”, “Sharpblue”, “Star”, “Suziblue”) and one rabbiteye cultivar (“Ochlockonee”). Plants were cultivated in soil with rows oriented east–west, and there were 15–18 plants of a given cultivar within a row. The experimental field was originally set up to assess the suitability of these cultivars for growth and production under the region's climatic conditions. Northern highbush and southern highbush blueberry differ in the number of chilling hours required for normal floral development, with northern highbush having higher chilling requirements (800 to 1000 h), southern highbush having lower chilling requirements (400 to 800 h) and rabbiteye requiring around 600 h (Retamales and Hancock, 2018). We randomly selected five plants per cultivar along the respective row to evaluate flower morphological traits and perform pollinator observations.
The experimental field was located in a landscape with multiple land-cover and land-use types, including farming systems (a mix of fruit orchards), grasslands, and forest and urban areas, similar to many production areas in Portugal. The study site was pollinator friendly (i.e. low disturbance, low inputs, and diverse floral and nesting resources). No beehives were added to the study site.
2.2 Flower morphological traits
Corolla length, width and aperture diameter were measured following the methodology of Courcelles et al. (2013), as these traits can potentially affect pollinator behaviour. Measurements were collected from 6–10 flowers for each of the five plants selected per cultivar. Flowers were collected into microtubes with 70 % ethanol and measured in the laboratory with a digital calliper. Briefly, we measured the corolla length by taking the flower's total length from the floral tube's base to the corolla's opening, the corolla's width at the widest point and the corolla opening diameter.
2.3 Pollinator assemblage
Floral visitors were observed at each cultivar's flowering peak on sunny days (temperature range 13–25 °C) with favourable weather conditions for pollination, from 15 March to 20 April 2023. Floral visits were recorded at different times of the day from 9:00 AM to 5:00 PM (CET+1 h), totalling 50 h 17 min of surveillance. The monitoring of floral visitors followed standard protocols (Dafni et al., 2005). Briefly, on each monitoring day, flowering plants were selected haphazardly and observed for 5 min periods. As several cultivars were simultaneously in flower, the observer alternated observations among cultivars after each 5 min period. Both the selection of individual plants and the sequence of cultivars were random. For each selected plant in each cultivar, we recorded the number and identity of individual insects that interacted with the flowers, insect behaviour (i.e. legitimate visit and primary or secondary nectar robber), and the cultivar and number of flowers visited. The interaction with each flower was considered a legitimate visit when the insect inserted its proboscis and/or head into the corolla or robbing when the insect positioned itself over the corolla, creating holes in its base (primary robber), or collecting nectar from existing holes (secondary robber). We used photographs and targeted captures to identify the insect visitors. Additionally, the total number of open flowers per cultivar was also recorded.
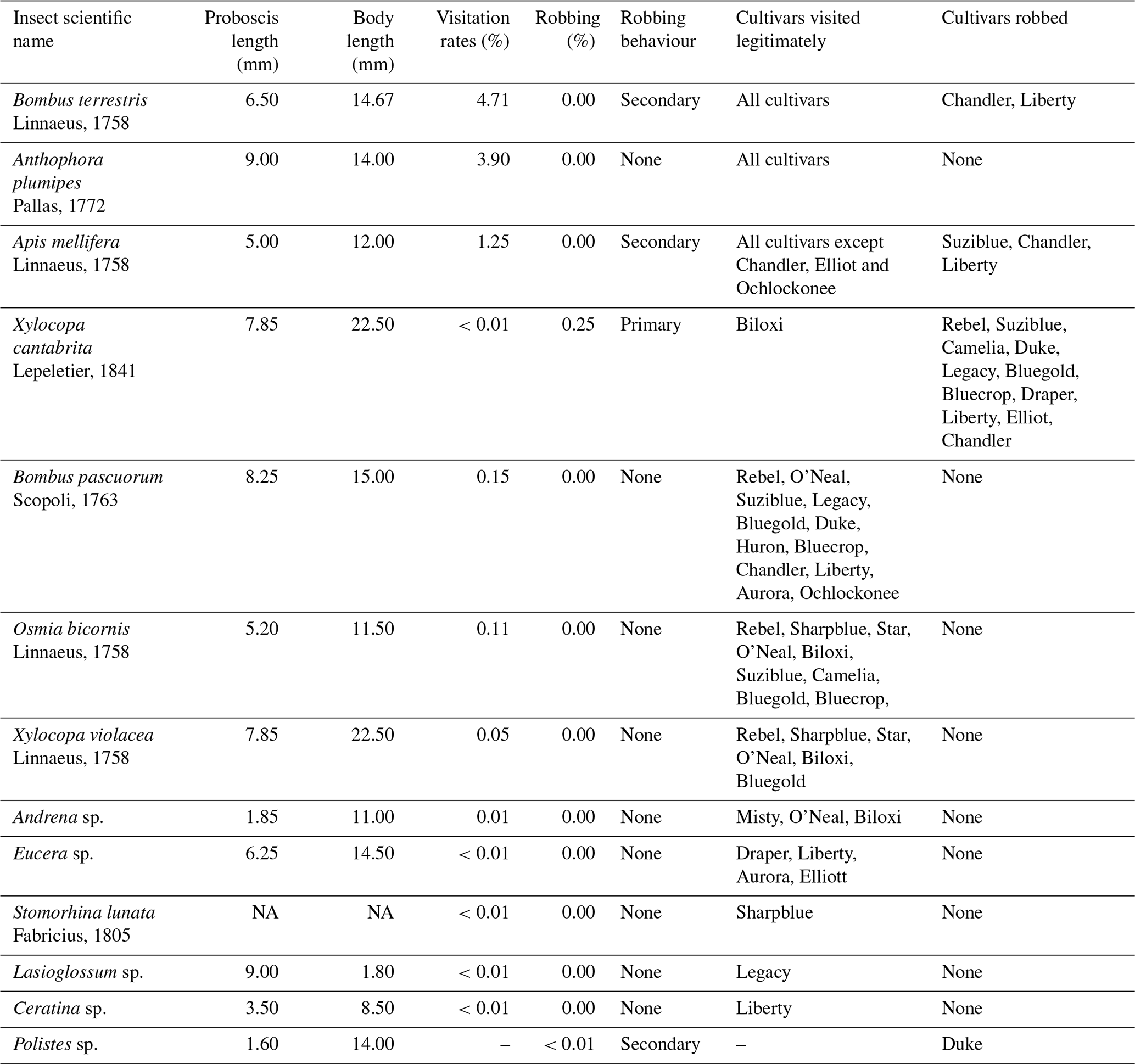
For each selected plant per cultivar, the visitation rates were calculated as the ratio of legitimately visited to open flowers per 5 min period. Values are presented as percentages of the flowers visited. In addition to calculating overall visitation rates, we computed the percentage of flowers visited by each of the three most common floral visitors (Bombus terrestris, Linnaeus, 1758; Anthophora plumipes, Pallas, 1772; and Apis mellifera) and the percentage of flowers robbed. For pollinator traits, we retrieved average values of body and proboscis length for these three species from Cappellari et al. (2022) to characterise the relative differences among these visitors.
Table 1Insect visitors recorded in the study interacting with blueberry flowers, their proboscis length (in mm), body length (in mm) and percentage of flowers visited by each species (visitation rate). The percentage of flowers robbed by each species (robbing), the type of robbing behaviour (primary or secondary) and the cultivars in which legitimate visits and robbing were recorded are also provided. Values for proboscis length and body length were obtained from Cappellari et al. (2022). NA – not available.
2.4 Data analyses
Flower morphological data and visitation rates were averaged per individual plant in each cultivar. To test for the effect of cultivar on flower morphology, visitation rates and robbing, we used general linear models with corolla length, corolla width, corolla opening diameter, visitation rates or percentage of flowers robbed as a dependent variable and cultivar as a fixed factor. We used principal component analysis to integrate the flower morphological traits, which are correlated to each other, using prcom, FactoMiner and facto extra (Kassambara and Mundt, 2020; Lê et al., 2008). The scores of the first PCA axis were extracted and used as a predictor to explore the effect of flower size on pollinator behaviour. The relationship between flower morphological traits and visitation rates by B. terrestris, An. plumipes and A. mellifera was assessed using generalised additive models (GAMs), with the visitation rate by each floral visitor as a dependent variable and the PCA first axis scores as a predictor (gam(visit_insect ∼ s(PC1), data = data, method = REML)). The mgcv package was used, and the basis dimensions and model fit were evaluated using gam.check. We used the ggplot package to build plots. Model fit was assessed using the diagnostic plots of the DHARMa package (Hartig, 2016). All analyses were performed using R software, version 4.3.0 (Core Development Team R, 2016).
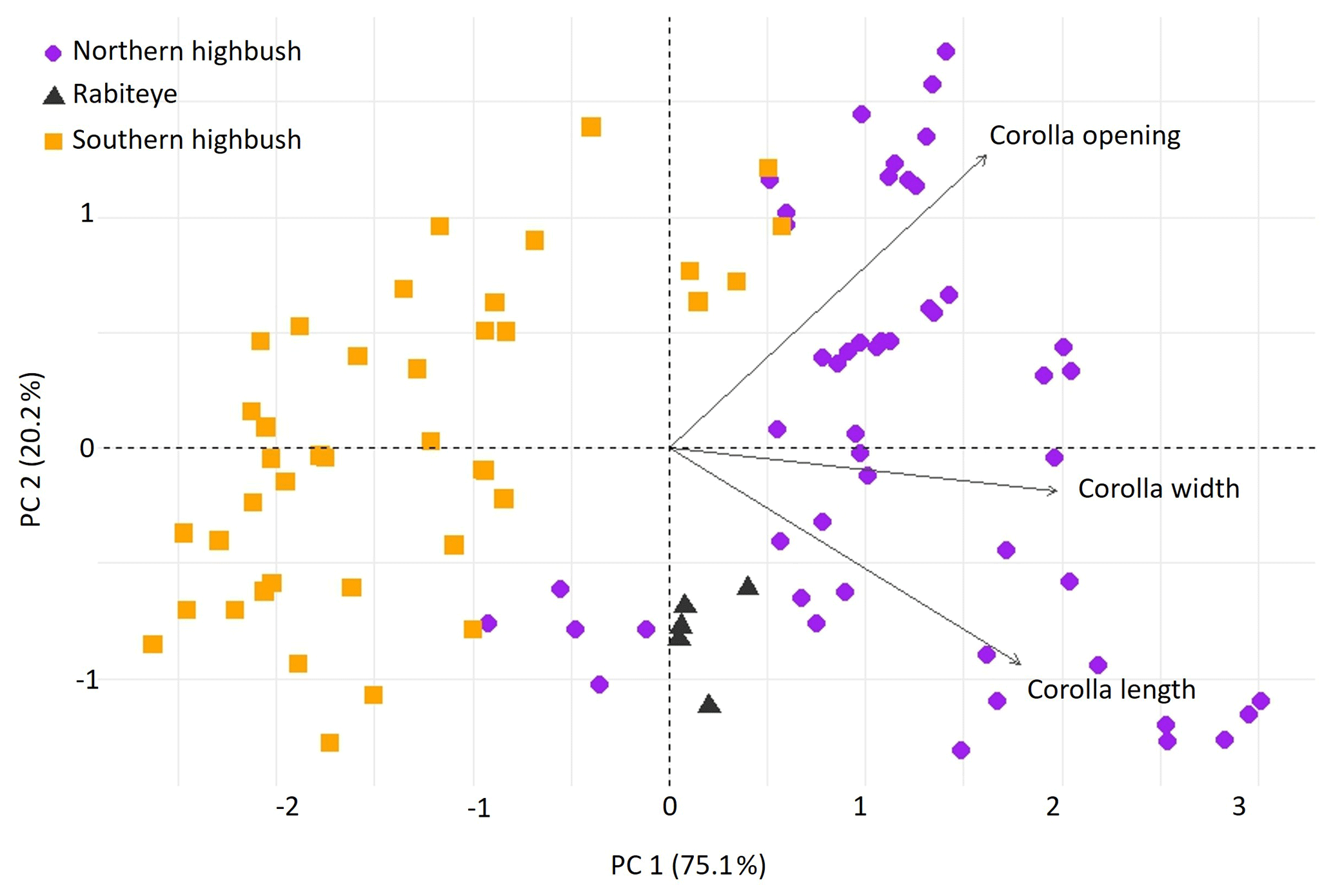
Figure 2Scatter plot of principal component analysis of flower morphological traits of 19 blueberry cultivars. Squares represent cultivars from the southern highbush group, dots represent the northern highbush group and triangles represent the cultivar from the rabbiteye group.
3.1 Flower morphological traits
The flower morphology of the studied blueberry cultivars at the study site differed significantly (Fig. 2, Supplement Fig. S1). Southern highbush cultivars tended to have smaller corollas than northern highbush cultivars, while the rabbiteye presented intermediate values (Fig. 2, Supplement Fig. S1). Corolla length ranged from 6.21 mm in “Star” to 11.85 mm in “Chandler”, while corolla width ranged from 4.18 mm in “Rebel” to 7.97 mm in “Draper”. Finally, the corolla opening ranged from 2.26 mm in “Rebel” to 5.13 mm in “Aurora” (Supplement Table S1 and Fig. S1).
3.2 Insect assemblage
A total of 13 insect species were observed interacting with the blueberry flowers (Table 1). Among these, three species accounted for 93.8 % of insect interactions with blueberry flowers, with the wild buff-tailed bumble bee (B. terrestris; proboscis 6.50 mm long) being the most frequent visitor (Supplement Fig. S2) with visitation rates at 4.71 %. The longer-tongued wild bee An. plumipes (proboscis 9.00 mm long) was the second most abundant floral visitor, followed by the shorter-tongued managed honeybee (A. mellifera; proboscis 5.00 mm long).
3.3 Effect of corolla morphological traits on plant–pollinator interactions
The percentage of flowers with legitimate visits differed significantly across cultivars (X18= 84.66, p < 0.0001; Fig. 3A). The lowest values were observed in “Chandler” (5.36 %), while the highest values were observed in “Aurora” (16.5 %). The percentage of flowers robbed also differed among cultivars (X18= 130.53, p < 0.0001; Fig. 3B), with “Chandler” showing the highest values (2.87 %).
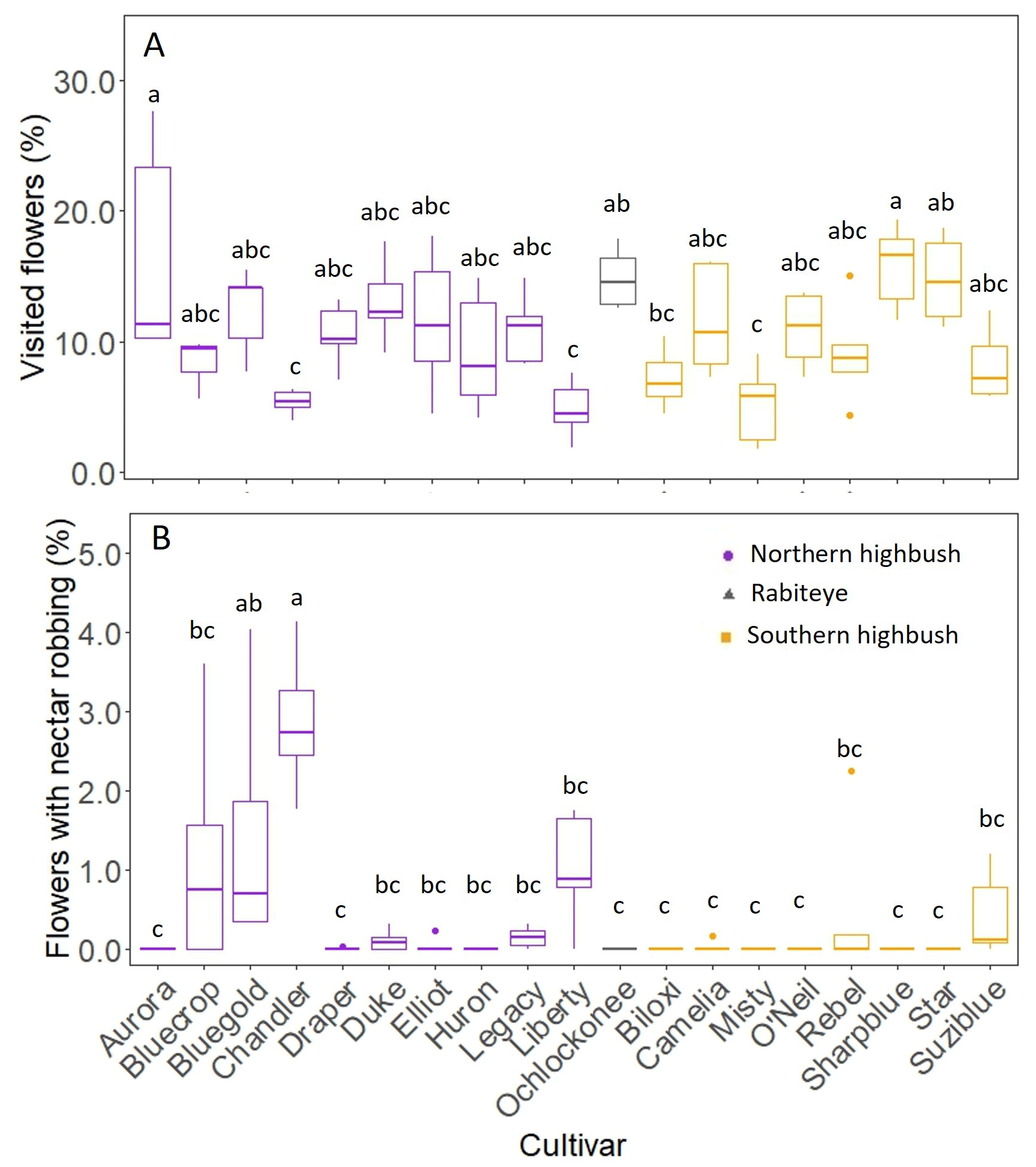
Figure 3Boxplot showing variation among 19 blueberry cultivars in the rate of legitimate floral visits (A) and the percentage of flower robbing (B). The line inside the box represents the median value, the box shows the interquartile range, the whiskers show the data range and the dots indicate outliers. Different letters indicate significant differences among cultivars at p<0.05.
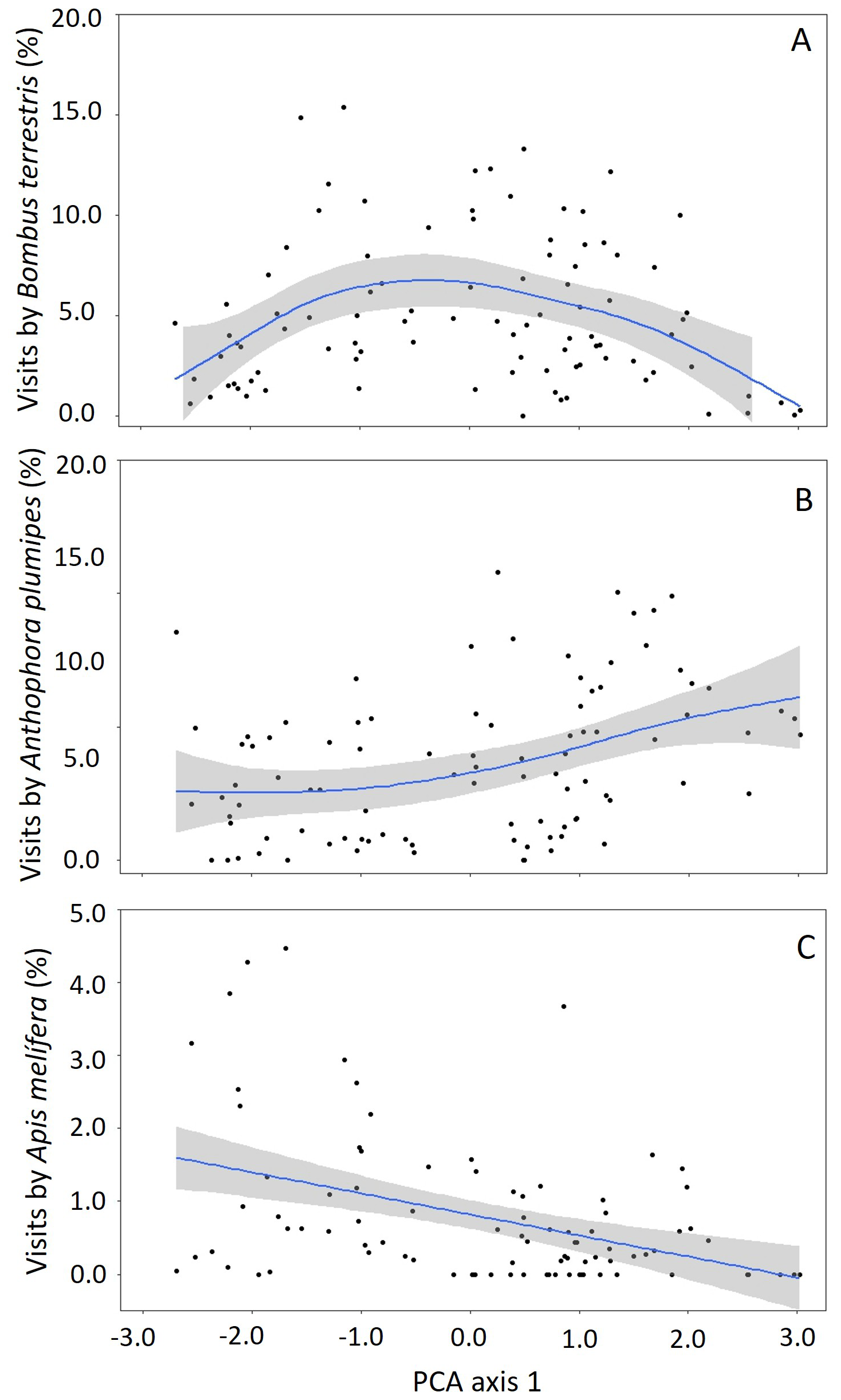
Figure 4Relationship between visitation rate by each floral visitor: Bombus terrestris (A), Anthophora plumipes (B) and Apis mellifera (C), along with the PCA first axis scores, representing a weighted combination of corolla length, width and opening. This provides a gradient in flower size where lower values in the PCA axis 1 represent smaller flowers.
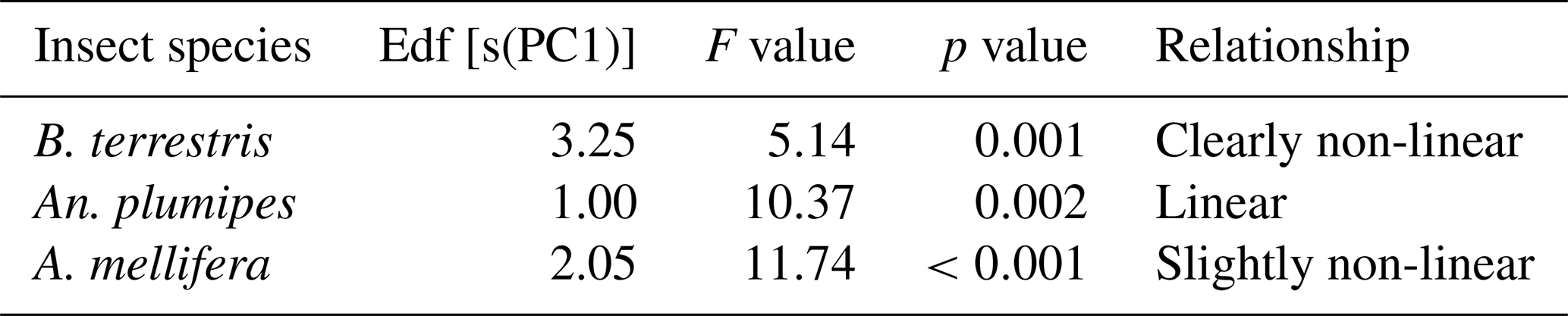
The results from the generalised additive models testing the effect of flower size – represented by the weighted combination of corolla length, width and opening (PCA axis 1) – on the behaviour of the main visitors of blueberry flowers show that floral visitors did not visit cultivars equally; B. terrestris had more visits to cultivars with intermediate corolla sizes (Fig. 4A; Table 2), An. plumipes visited more flowers on cultivars with large corollas (Fig. 4B; Table 2) and A. mellifera tended to visit cultivars with small corollas (Fig. 4C; Table 2).
This study aimed to explore the impact of blueberry floral traits on pollinator visitation and behaviour. Blueberry cultivars differed in flower morphology, which is consistent with the findings from other studies (e.g. Arrington and DeVetter, 2018; Courcelles et al., 2013; Ritzinger and Lyrene, 1999; Lyrene, 1994). We found considerable variation in corolla length associated with variation in corolla width and corolla opening, leading to a gradient in flower size. The variation in corolla morphology influenced the identity and behaviour of floral visitors. We identified three main floral visitors: A. mellifera, which tended to visit cultivars with smaller flowers; An. plumipes, which tended to visit cultivars with larger flowers; and B. terrestris, which visited cultivars with flowers of intermediate size. The results are discussed in detail below.
Table 2Results from the generalised additive models testing the effect of flower size on the behaviour of the main visitors of blueberry flowers at the study site. Effective degrees of freedom (Edf) indicating linear or non-linear relationship (values >2 indicate non-linear relationship), F value and p value are reported.
4.1 Effect of corolla morphological traits on pollinators
Some studies indicate that a diverse assemblage of insects contributes to blueberry pollination, with honeybees showing the most significant abundance, followed by bumblebees and solitary bees (e.g. Cortés-Rivas et al., 2023b; Eeraerts et al., 2023). This pattern was also observed by Isaacs and Kirk (2010) on large commercial farms, but not on small farms, where they found that wild bees were the primary pollinators, and A. mellifera was less common than wild bees. In our study, B. terrestris and An. plumipes were the main visitors, followed by A. mellifera, while other wild bees were observed interacting at very low frequencies. Consistent with our findings, Cane and Payne (1993) report the predominance of a few species visiting rabbiteye blueberries in the southeastern United States, including honeybees, bumblebees and the southeastern blueberry bee (Habropoda laboriosa). Our study site is more closely aligned with the characteristics of small farms described by Isaacs and Kirk (2010) and Cane and Payne (1993), and is likely to provide areas with suitable habitats for diverse and abundant floral resources and nesting sites, contributing to a greater abundance of wild bees. The low abundance of honeybees could be partially due to the absence of beehives at the study site, unlike the standard practice in commercial orchards, and to the proximity of an apple orchard, which overlapped in flowering with blueberry and may be more attractive to honeybees due to easier access to resources offered by the open flower morphology.
Flowers such as those of the blueberry, with morphological traits that condition access to pollen and rewards, will affect pollinator visitation and behaviour. Indeed, our data support this, as we found distinct relationships between the visitation rates of each of the three main floral visitors and flower size. Bombus terrestris visited more flowers on cultivars with intermediate flower sizes, An. plumipes visited more cultivars with larger flowers and A. mellifera tended to visit cultivars with smaller flowers. These differences may be linked to the relationship between proboscis size and flower morphology, which limits access to rewards and determines pollinator behaviour (Klumpers et al., 2019). For example, bumblebee species that differ in proboscis length exhibit differences in flower visitation across a range of flower-tube lengths (Inouye, 1980). This may be related to flower handling time and efficiency in nectar extraction, where pollinators with proboscides shorter than the corolla require more time to collect nectar compared to pollinators with proboscides that match corolla size (Klumpers et al., 2019). Higher handling efficiency implies a lower cost-benefit in foraging and higher fitness, leading pollinators to prefer flowers they can handle more efficiently (Balfour et al., 2013). The average proboscis lengths of the main pollinators of blueberry may explain the patterns observed in our study. Apis mellifera, with the shortest proboscis (5.00 mm) of the three species, visited cultivars with smaller flowers; An. plumipes, with the longest proboscis (9.00 mm), visited cultivars with larger flowers; and B. terrestris, with intermediate values (6.5 mm), visited cultivars with intermediate sizes. While bees with long proboscides can visit both flowers with long and short corollas, there is evidence indicating that the handling time and the number of flowers visited per unit of time are positively correlated to proboscis length (Balfour et al., 2013; Herrera, 1989; Inouye, 1980). For example, proboscis length and resource extraction time were negatively correlated in lavender, and when the length of the corolla was experimentally reduced, the handling time by A. mellifera decreased (Balfour et al., 2013). Results suggest that the three main visitors of blueberry in our study select flowers that best fit their morphology, allowing them to forage more efficiently, and show evidence of resource partitioning, where An. plumipes visits flowers less accessible to and/or less efficiently handled by other visitors with a shorter proboscis. Nevertheless, other factors that influence insect behaviour, such as nectar volume and sugar content (Klumpers et al., 2019), have not been assessed, and their contributions need to be considered in future studies.
We observed primary nectar robbing by the carpenter bee Xylocopa cantabrita Lepeletier, 1841 and secondary robbing by B. terrestris and A. mellifera. Similar behaviours were also reported by Cane and Payne (1993) and Sampson et al. (2004) for X. virginica Linnaeus, 1771 (absent in Europe) and A. mellifera, where the carpenter bee pierces the base of the corolla, and honeybees profit from the greater accessibility to the nectaries and engage in secondary nectar robbing. Nectar robbing can influence the patterns of nectar availability and consequently affects the attractiveness of the flower to pollinators, resulting in lower rates of legitimate visits and lower fruit production (Castro et al., 2008). Nevertheless, despite this behaviour, carpenter bees are reported to deposit pollen tetrads on the stigma on their flower visits, thereby contributing to blueberry pollination, particularly in cultivars with protruding stigmas (Sampson et al., 2004; Tucker et al., 2019).
4.2 Implications for management
Overall, our findings indicate that a variation in flower morphological traits across blueberry cultivars influences pollinator visitation rates, with proboscis length likely linked to differences in nectar foraging preferences among the evaluated taxa. Breeding programmes are encouraged to consider flower traits and their influence on pollination success in commercial settings. Likewise, growers should select cultivars that can be readily pollinated by the insect communities they rely on for pollination services, given that pollination success is a prerequisite for commercially viable yields and fruit quality. Moreover, promoting wild pollinators by maintaining or creating natural areas with flower and nesting resources increases the probability of having species with different proboscis sizes, ensuring legitimate visits to a wider range of cultivars, as well as species with the capacity for buzz pollination (e.g. Bombus spp., Anthophora spp.), which are more efficient at pollinating flowers with poricidal anthers (Buchmann and Hurley, 1978; De Luca et al., 2019), such as those of the blueberry. Conversely, if the honeybee is to be the primary pollinator, care should be taken to select cultivars with shorter and/or wider corollas, allowing the honeybees to reach nectar by inserting their heads inside the corolla, preferably with extruding pistils that allow contact with honeybee body parts carrying pollen (Hoffman et al., 2018).
This study demonstrates that blueberry cultivars differ markedly in flower morphology and that this variation has clear consequences for plant–pollinator interactions. Visitation rates and behavioural patterns of the main floral visitors were closely associated with corolla traits, supporting the hypothesis that longer corollas and smaller apertures impose greater constraints on floral visitors, while shorter and wider corollas are accessible to a broader assemblage of insects. Differences in pollinators' proboscis lengths were reflected in cultivar-specific visitation patterns, indicating functional matching between floral morphology and pollinator traits, and suggesting resource partitioning among visitors. In addition, cultivars with more restrictive floral morphologies were associated with higher levels of nectar robbing, highlighting a potential trade-off between floral specialisation and effective pollination. Together, these results indicate that both cultivar identity and pollinator assemblage composition are likely to influence blueberry pollination outcomes through their effects on pollinator visitation and behaviour. From an applied perspective, our findings also emphasise that flower morphology should be considered in blueberry breeding programmes and orchard design, as it influences plant–pollinator interactions and pollination success. Promoting diverse wild pollinator communities and strategically selecting cultivar combinations may therefore improve pollination services and contribute to more resilient and productive blueberry agroecosystems.
Data are available in the Figshare Repository at https://doi.org/10.6084/m9.figshare.32325837 (Castro et al., 2026).
The supplement related to this article is available online at https://doi.org/10.5194/we-26-83-2026-supplement.
HC designed the study, collected and analysed data, and produced the first draft of the paper. AMB, BC and HG collected data and contributed to the preparation of the paper. JL and SL contributed to the discussion of study design and preparation of the paper.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We thank Estação Agrária de Viseu – DRAPC for allowing us to work on their grounds.
HC was funded by national funds (OE) through Fundação para a Ciência e Tecnologia, I.P., within the scope of the framework contract foreseen in numbers 4–6 of article 23 of the Decree-Law 57/2016, 29 August, as amended by Law 57/2017, 19 July. This work was carried out at the R&D Unit Centre for Functional Ecology – Science for People and the Planet (CFE), with reference UIDB/04004/2020 and financed by FCT/MCTES through national funds (https://doi.org/10.54499/UIDB/04004/2020). HG was funded by the Integrated Program of Scientific Research and Technological Development CULTIVAR (CENTRO-01-0145-FEDER-000020), co-financed by the Regional Operational Programme Centro 2020, Portugal 2020 and European Union, through the European Fund for Regional Development (ERDF) and by the Portuguese Foundation for Science and Technology (FCT – Fundação para a Ciência e a Tecnologia, I.P.) through the fellowship 2023.01736.BD.
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
This paper was edited by Carine Emer and reviewed by Laura Stefan and three anonymous referees.
Arrington, M. and DeVetter, L. W.: Floral Morphology Differs Among New Northern Highbush Blueberry Cultivars, J. Hortic., 05, 10–13, https://doi.org/10.4172/2376-0354.1000223, 2018.
Balfour, N. J., Garbuzov, M., and Ratnieks, F. L. W.: Longer tongues and swifter handling: Why do more bumble bees (Bombus spp.) than honey bees (Apis mellifera) forage on lavender (Lavandula spp.)?, Ecol. Entomol., 38, 323–329, https://doi.org/10.1111/een.12019, 2013.
Buchmann, S. L. and Hurley, J. P.: A biophysical model for buzz pollination in angiosperms, J. Theor. Biol., 72, 639–657, https://doi.org/10.1016/0022-5193(78)90277-1, 1978.
Cane, J. H. and Payne, J. A.: Regional, Annual, and Seasonal Variation in Pollinator Guilds: Intrinsic Traits of Bees (Hymenoptera: Apoidea) Underlie Their Patterns of Abundance at Vaccinium ashei (Ericaceae), Ecol. Popul. Biol., 86, 577–588, 1993.
Cane, James H. and Payne, Jerry A.: Foraging Ecology of the Bee Habropoda laboriosa (Hymenoptera: Anthophoridae), an Oligolege of Blueberries (Ericaceae: Vaccinium) in the Southeastern United States, Ann. Entomol. Soc. Am., 75, 419–427, 1988.
Cappellari, A., Bonaldi, G., Mei, M., Paniccia, D., Cerretti, P., and Marini, L.: Functional traits of plants and pollinators explain resource overlap between honeybees and wild pollinators, Oecologia, 198, 1019–1029, https://doi.org/10.1007/s00442-022-05151-6, 2022.
Castro, H., Siopa, C., Casais, V., Castro, M., Loureiro, J., Gaspar, H., Dias, M. C., and Castro, S.: Spatiotemporal variation in pollination deficits in an insect-pollinated dioecious crop, Plants, 10, https://doi.org/10.3390/plants10071273, 2021.
Castro, H., Branco, A. M., Chaves, H., Gaspar, H., Loureiro, J., and Castro, S.: Blueberry floral morphology influences pollinator visitation and behavioural patterns, Figshare Repository [code], https://doi.org/10.6084/m9.figshare.32325837, 2026.
Castro, S., Silveira, P., and Navarro, L.: Consequences of nectar robbing for the fitness of a threatened plant species, Plant Ecol., 199, 201–208, https://doi.org/10.1007/s11258-008-9424-z, 2008.
Core Development Team R: A language and environment for statistical computing, Vienna: R Foundation for Statistical Computing, https://www.R-project.org/ (last access: 4 June 2026), 2016.
Cortés-Rivas, B., Monzón, V. H., Rego, J. O., and Mesquita-Neto, J. N.: Pollination by native bees achieves high fruit quantity and quality of highbush blueberry: a sustainable alternative to managed pollinators, Front. Sustain. Food Syst., 7, https://doi.org/10.3389/fsufs.2023.1142623, 2023a.
Cortés-Rivas, B., Smith-Ramirez, C., Monzón, V. H., and Mesquita-Neto, J. N.: Native bees with floral sonication behaviour can achieve high-performance pollination of highbush blueberry in Chile, Agric. For. Entomol., 25, 91–102, https://doi.org/10.1111/afe.12533, 2023b.
Courcelles, D. M. M., Button, L., and Elle, E.: Bee visit rates vary with floral morphology among highbush blueberry cultivars (Vaccinium corymbosum L.), J. Appl. Entomol., 137, 693–701, https://doi.org/10.1111/jen.12059, 2013.
Dafni, A., Kevan, P. G., and Husband, B. C.: Practical Pollination Ecology, Enviroquest, Ltd., 590 pp., ISBN 0-19-963298-7, 2005.
De Luca, P. A. and Vallejo-Marín, M.: What's the “buzz” about? The ecology and evolutionary significance of buzz-pollination, Curr. Opin. Plant Biol., 16, 429–435, https://doi.org/10.1016/j.pbi.2013.05.002, 2013.
De Luca, P. A., Buchmann, S., Galen, C., Mason, A. C., and Vallejo-Marín, M.: Does body size predict the buzz-pollination frequencies used by bees?, Ecol. Evol., 9, 4875–4887, https://doi.org/10.1002/ece3.5092, 2019.
DeVetter, L. W., Chabert, S., Milbrath, M. O., Mallinger, R. E., Walters, J., Isaacs, R., Galinato, S. P., Kogan, C., Brouwer, K., Melathopoulos, A., and Eeraerts, M.: Toward evidence-based decision support systems to optimize pollination and yields in highbush blueberry, Front. Sustain. Food Syst., 6, https://doi.org/10.3389/fsufs.2022.1006201, 2022.
Dogterom, M. H., Winston, M. L., and Mukai, A.: Effect of pollen load size and source (self, outcross) on seed and fruit production in highbush blueberry cv. “Bluecrop” (VACCINIUM CORYMBOSUM; Ericaceae), Am. J. Bot., 87, 1584–1591, https://doi.org/10.2307/2656734, 2000.
Eck, P. and Mainland, C. M.: Highbush Blueberry Fruit Set in Relation to Flower Morphology, HortScience, 6, 494–495, 1971.
Edger, P. P., Iorizzo, M., Bassil, N. V., Benevenuto, J., Ferrão, L. F. V., Giongo, L., Hummer, K., Lawas, L. M. F., Leisner, C. P., Li, C., Munoz, P. R., Ashrafi, H., Atucha, A., Babiker, E. M., Canales, E., Chagne, D., Devetter, L., Ehlenfeldt, M., Espley, R. V., Gallardo, K., Günther, C. S., Hardigan, M., Hulse-Kemp, A. M., Jacobs, M., Lila, M. A., Luby, C., Main, D., Mengist, M. F., Owens, G. L., Perkins-Veazie, P., Polashock, J., Pottorff, M., Rowland, L. J., Sims, C. A., Song, G. Q., Spencer, J., Vorsa, N., Yocca, A. E., and Zalapa, J.: There and back again; historical perspective and future directions for Vaccinium breeding and research studies, Hortic. Res., 9, https://doi.org/10.1093/hr/uhac083, 2022.
Eeraerts, M., DeVetter, L. W., Batáry, P., Ternest, J. J., Mallinger, R., Arrington, M., Benjamin, F. E., Blaauw, B. R., Campbell, J. W., Cavigliasso, P., Daniels, J. C., de Groot, G. A., Ellis, J. D., Gibbs, J., Goldstein, L., Hoffman, G. D., Kleijn, D., Melathopoulos, A., Miller, S. Z., Montero-Castaño, A., Naranjo, S. M., Nicholson, C. C., Perkins, J. A., Rao, S., Raine, N. E., Reilly, J. R., Ricketts, T. H., Rogers, E., and Isaacs, R.: Synthesis of highbush blueberry pollination research reveals region-specific differences in the contributions of honeybees and wild bees, J. Appl. Ecol., 60, 2528–2539, https://doi.org/10.1111/1365-2664.14516, 2023.
Ehlenfeldt, M. K.: Self- and cross-fertility in recently released highbush blueberry cultivars, HortScience, 36, 133–135, https://doi.org/10.21273/hortsci.36.1.133, 2001.
Gérard, M., Vanderplanck, M., Wood, T., and Michez, D.: Global warming and plant-pollinator mismatches, Emerg. Top. Life Sci., 4, 77–86, https://doi.org/10.1042/ETLS20190139, 2020.
Hartig, F.: DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models, GitHub [data set], https://github.com/florianhartig/dharma (last access: 4 June 2026), 2016.
Herrera, C. M.: Pollinator Abundance, Morphology, and Flower Visitation Rate: Analysis of the “Quantity” Component in a Plant-Pollinator System, Oecologia, 80, 241–248, https://doi.org/10.1007/bf00380158, 1989.
Hoffman, G. D., Lande, C., and Rao, S.: A novel pollen transfer mechanism by honey bee foragers on highbush blueberry (Ericales: Ericaceae), Environ. Entomol., 47, 1465–1470, https://doi.org/10.1093/ee/nvy162, 2018.
Inouye, D. W.: The effect of proboscis and corolla tube lengths on patterns and rates of flower visitation by bumblebees, Oecologia, 45, 197–201, https://doi.org/10.1007/BF00346460, 1980.
Irwin, R. E., Bronstein, J. L., Manson, J. S., and Richardson, L.: Nectar robbing: Ecological and evolutionary perspectives, Annu. Rev. Ecol. Evol. Syst., 41, 271–292, https://doi.org/10.1146/annurev.ecolsys.110308.120330, 2010.
Isaacs, R. and Kirk, A. K.: Pollination services provided to small and large highbush blueberry fields by wild and managed bees, J. Appl. Ecol., 47, 841–849, https://doi.org/10.1111/j.1365-2664.2010.01823.x, 2010.
Kassambara, A. and Mundt, F.: Factoextra: Extract and Visualize the Results of Multivariate Data Analyses, R package version 2.0.0 [code], https://CRAN.R-project.org/package=factoextra (last access: 4 June 2026), 2020.
Kendall, L. K., Gagic, V., Evans, L. J., Cutting, B. T., Scalzo, J., Hanusch, Y., Jones, J., Rocchetti, M., Sonter, C., Keir, M., and Rader, R.: Self-compatible blueberry cultivars require fewer floral visits to maximize fruit production than a partially self-incompatible cultivar, J. Appl. Ecol., 57, 2454–2462, https://doi.org/10.1111/1365-2664.13751, 2020.
Kendall, L. K., Stavert, J. R., Gagic, V., Hall, M., and Rader, R.: Initial floral visitor identity and foraging time strongly influence blueberry reproductive success, Basic Appl. Ecol., 60, 114–122, https://doi.org/10.1016/j.baae.2022.02.009, 2022.
Klumpers, S. G. T., Stang, M., and Klinkhamer, P. G. L.: Foraging efficiency and size matching in a plant–pollinator community: the importance of sugar content and tongue length, Ecol. Lett., 22, 469–479, https://doi.org/10.1111/ele.13204, 2019.
Kumarihami, H. M. P. C., Park, H. G., Kim, S. M., Park, J. I., Lee, E. J., Kim, H. L., and Kim, J. G.: Flower and leaf bud density manipulation affects fruit set, leaf-to-fruit ratio, and yield in southern highbush “Misty” blueberry, Sci. Hortic. (Amsterdam), 290, 110530, https://doi.org/10.1016/j.scienta.2021.110530, 2021.
Kuppler, J. and Kotowska, M. M.: A meta-analysis of responses in floral traits and flower–visitor interactions to water deficit, Glob. Chang. Biol., 27, 3095–3108, https://doi.org/10.1111/gcb.15621, 2021.
Lê, S., Josse, J., and Husson, F.: FactoMineR: An R Package for Multivariate Analysis, 2008.
Lyrene, P. M.: Variation within and among blueberry taxa in flower size and shape, J. Am. Soc. Hortic. Sci., 119, 1039–1042, https://doi.org/10.21273/jashs.119.5.1039, 1994.
Nicholson, C. C., Koh, I., Richardson, L. L., Beauchemin, A., and Ricketts, T. H.: Farm and landscape factors interact to affect the supply of pollination services, Agric. Ecosyst. Environ., 250, 113–122, https://doi.org/10.1016/j.agee.2017.08.030, 2017.
Payne, J. A., Amis, A. A., Cane, J. H., and Lyrene, P. M.: Fruit size, seed size, seed viability and pollination of rabbiteye blueberries (Vaccinium ashei), Acta Hortic., 38–43, https://doi.org/10.17660/ActaHortic.1989.241.3, 1989.
Porto, R. G., de Almeida, R. F., Cruz-Neto, O., Tabarelli, M., Viana, B. F., Peres, C. A., and Lopes, A. V.: Pollination ecosystem services: A comprehensive review of economic values, research funding and policy actions, Food Secur., 12, 1425–1442, https://doi.org/10.1007/s12571-020-01043-w, 2020.
Potts, S. G., Biesmeijer, J. C., Kremen, C., Neumann, P., Schweiger, O., and Kunin, W. E.: Global pollinator declines: Trends, impacts and drivers, Trends Ecol. Evol., 25, 345–353, https://doi.org/10.1016/j.tree.2010.01.007, 2010.
Retamales, J. and Hancock, J. F.: Blueberries, 2nd Edition., CABI, 411 pp., ISBN 978-1-78064-726-5, 2018.
Ritzinger, R. and Lyrene, P. M.: Flower morphology in blueberry species and hybrids, HortScience, 34, 130–131, https://doi.org/10.21273/hortsci.34.1.130, 1999.
Sampson, B. J., Danka, R. G., and Stringer, S. J.: Nectar robbery by bees Xylocopa virginica and Apis mellifera contributes to the pollination of rabbiteye blueberry, J. Econ. Entomol., 97, 735–740, https://doi.org/10.1093/jee/97.3.735, 2004.
Sampson, B. J., Stringer, S. J., and Marshall, D. A.: Blueberry floral attributes and their effect on the pollination efficiency of an oligolectic bee, Osmia ribifloris Cockerell (Megachilidae: Apoidea), HortScience, 48, 136–142, https://doi.org/10.21273/hortsci.48.2.136, 2013.
Song, G.-Q. and Hancock, J. F.: Vaccinium, in: Wild Crop Relatives: Genomic and Breeding Resources, edited by: Kole, C., Springer Berlin Heidelberg, Berlin, Heidelberg, 197–221, https://doi.org/10.1007/978-3-642-16057-8_10, 2011.
Stang, M., Klinkhamer, P. G. L., Waser, N. M., Stang, I., and Van Der Meijden, E.: Size-specific interaction patterns and size matching in a plant-pollinator interaction web, Ann. Bot., 103, 1459–1469, https://doi.org/10.1093/aob/mcp027, 2009.
Symington, H. A. and Glover, B. J.: Strawberry varieties differ in pollinator-relevant floral traits, Ecol. Evol., 14, 1–14, https://doi.org/10.1002/ece3.10914, 2024.
Taber, S. K. and Olmstead, J. W.: Impact of cross- and self-pollination on fruit set, fruit size, seed number, and harvest timing among 13 southern highbush blueberry cultivars, Horttechnology, 26, 213–219, https://doi.org/10.21273/horttech.26.2.213, 2016.
Tucker, S. K., Ginsberg, H. S., and Alm, S. R.: Effect of corolla slitting and nectar robbery by the Eastern Carpenter Bee (Hymenoptera: Apidae) on fruit quality of Vaccinium corymbosum L. (Ericales: Ericaceae), Environ. Entomol., 48, 718–726, https://doi.org/10.1093/ee/nvz055, 2019.
Vander Kloet, S. P.: The Genus Vaccinium, 201 pp., Canadian Government Publishing Centre, Ottawa, 1988.
Woodcock, B. A., Garratt, M. P. D., Powney, G. D., Shaw, R. F., Osborne, J. L., Soroka, J., Lindström, S. A. M., Stanley, D., Ouvrard, P., Edwards, M. E., Jauker, F., McCracken, M. E., Zou, Y., Potts, S. G., Rundlöf, M., Noriega, J. A., Greenop, A., Smith, H. G., Bommarco, R., van der Werf, W., Stout, J. C., Steffan-Dewenter, I., Morandin, L., Bullock, J. M., and Pywell, R. F.: Meta-analysis reveals that pollinator functional diversity and abundance enhance crop pollination and yield, Nat. Commun., 10, https://doi.org/10.1038/s41467-019-09393-6, 2019.