the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 18 Jun 2026
| 18 Jun 2026
Invisible extinction in a sacred refuge: a dying micro-population of Glyptostrobus pensilis at Nanhua Temple, China
Chunping Xie
Dawei Liu
Extremely small and isolated remnant populations represent an overlooked but critical conservation challenge for many threatened plant species. Although such populations may still be recorded as extant, demographic collapse and regeneration failure often place them at high risk of local extinction. Here we report the rapid decline of a micro-population of Glyptostrobus pensilis, a globally threatened relict conifer, discovered in a sacred forest associated with the millennium-old Nanhua Temple in southern China. Field observations identified only nine mature individuals, all of which were centuries old and showed signs of physiological deterioration. At least two trees had already died, and no seedlings or saplings were observed, indicating long-term recruitment failure and demographic non-viability. Despite its location within a culturally protected landscape, the population lacks ecological management and habitat restoration. This case highlights the limitations of passive protection and illustrates the risk of “invisible extinction” in sacred or cultural refugia. We argue that micro-populations should be recognized as high-priority conservation units and recommend the establishment of monitoring thresholds, emergency demographic interventions, habitat restoration, and integration of cultural landscapes into science-based conservation strategies.
- Article
(3416 KB) - Full-text XML
- BibTeX
- EndNote




Global conservation paradigms for threatened flora have traditionally been anchored by two predominant metrics: geographical range extent and aggregate population abundance (Nic Lughadha et al., 2020; Yang et al., 2020). These parameters constitute the bedrock of major evaluative frameworks, most notably the IUCN Red List, which relies on quantitative thresholds of population size and range contraction to delineate extinction risk (Rønsted et al., 2022). While these standardized metrics are indispensable for macro-level conservation prioritization, they harbor a critical methodological blind spot: they frequently overlook the precarious status of remnant micro-populations. Such populations, characterized by extreme numerical depletion, spatial fragmentation, and functional senescence, may remain formally classified as “extant” while undergoing an irreversible trajectory toward “invisible extinction” (Crist, 2022). This discrepancy between taxonomic presence and demographic viability masks an accelerating collapse that standard monitoring protocols often fail to capture.
Across global biodiversity hotspots, threatened woody species increasingly persist as scattered micro-populations instead of robust metapopulations. These relictual assemblages often comprise fewer than a few dozen individuals (Tang et al., 2019) and are frequently confined to fragmented matrices such as secondary vegetation, riparian corridors, and anthropogenic refugia like culturally protected landscapes (Frascaroli et al., 2016; Samways et al., 2025). While these sites provide a temporary buffer against land-use conversion, such extreme isolation often severs the ecological processes necessary for long-term population persistence (Ensslin and Godefroid, 2019). These populations share a constellation of vulnerabilities that are rarely captured by conventional assessments. The extinction risk of micro-populations is primarily exacerbated by demographic stochasticity, where random fluctuations in vital rates significantly elevate the probability of collapse regardless of environmental stability (Lande, 1993). This vulnerability is further compounded by recruitment failure (Maschinski et al., 2023), often originating from disrupted dispersal vectors, altered hydrological regimes, or competitive exclusions. Such reproductive stagnation ensures that even long-lived individuals fail to produce a viable next generation. Furthermore, genetic erosion through inbreeding depression and the depletion of adaptive variation progressively compromises individual fitness and long-term population resilience (Leroy et al., 2018). Together, these processes can precipitate what has been termed an “extinction vortex” (Nordstrom et al., 2023; Gilpin and Soulé, 1986), in which mutually reinforcing demographic and genetic pressures drive a local population irreversibly toward zero.
The conservation science community has increasingly recognized the concept of functionally extinct populations, defined as biological assemblages that persist in situ yet fail to fulfill ecological roles or sustain self-replacement (Ledford, 2019). Yet institutional conservation responses lag behind this conceptual advance. Species-level threat assessments rarely disaggregate population-level viability (Mastretta-Yanes et al., 2024), and management resources are seldom directed at micro-populations. The result is a category of conservation urgency that is simultaneously high-risk and low-visibility: populations in silent decline, embedded in landscapes that appear protected, approaching extinction without triggering conservation alarm.
This problem is particularly acute in East and Southeast Asia, where rapid land-use change over the past century has fragmented the ranges of numerous endemic and relict tree species (Zheng et al., 2021; Li and Xia, 2004). Paradoxically, many of the final survivors of these species are located outside formal nature reserves. These individuals often persist within culturally protected spaces such as temple groves, village fengshui forests, and monastery grounds, where religious significance has historically afforded a degree of inadvertent protection (Zhou et al., 2022; Frascaroli et al., 2016; Sharma and Kumar, 2021; Xie et al., 2025). While the conservation value of such cultural refugia is well-documented (Huang et al., 2023), the extent to which they can sustain long-term population viability in the absence of active ecological management has received far less scrutiny. Effectively addressing this gap requires a multifaceted understanding of both the social and biological dimensions of sacred sites. While recent research has begun to bridge the divide between cultural reverence and scientific care through anthropological and mixed-methods approaches (Xie and Jim, 2026), there remains an urgent need for high-resolution demographic diagnostics to quantify the immediate extinction risks facing these relict populations.
We report the rapid demographic decline of a remnant micro-population of Glyptostrobus pensilis (Staunton ex D. Don) K. Koch, a globally threatened relict conifer located within the sacred forest of Nanhua Temple in Shaoguan, southern China (Li et al., 2004). This case epitomizes the phenomenon of invisible extinction within cultural refugia, highlighting a critical need to reframe conservation priorities toward population viability instead of mere taxonomic presence. We therefore argue that micro-populations warrant dedicated monitoring protocols, emergency interventions, and formal integration into conservation frameworks.
2.1 Species background
G. pensilis, widely referred to as the Chinese swamp cypress or water pine (shui song), represents one of the most ancient and evolutionarily distinct coniferous lineages in Asia (Hao et al., 2016). As the sole extant species of the monotypic genus Glyptostrobus, it represents an isolated lineage whose fossil record extends to the Cretaceous (Ben, 2007). During the Tertiary, the genus was distributed across the Northern Hemisphere, but its range contracted dramatically through the Quaternary glaciations, leaving G. pensilis as a glacial relict confined to subtropical China, northern Vietnam, and Laos (Li and Xia, 2004; Ye et al., 2022). The species is currently listed as Critically Endangered (CR) on the IUCN Red List and as a Category I State Key Protected Wild Plant in China, reflecting both its evolutionary irreplaceability and its precarious conservation status (Thomas et al., 2020; Yang et al., 2021).
G. pensilis is a deciduous, semi-aquatic conifer naturally adapted to riparian and waterlogged habitats, including river margins, swamp edges, and valley floors with seasonally fluctuating water tables (Tang et al., 2019). It develops distinctive pneumatophores (respiratory roots) under waterlogged conditions and is highly sensitive to hydrological modification. Historically, the species was more widely cultivated and naturalized across southeastern China, but the combined effects of wetland drainage, river channelization, land conversion, and over-harvesting have caused severe range contraction over the past century (Li and Xia, 2004; Li et al., 2004). Today, truly wild or semi-wild populations are highly fragmented, and many consist of only a handful of elderly individuals (Li et al., 2004; Yang et al., 2024). Despite its formal protected status, the species continues to decline across much of its residual range, and successful natural regeneration has become exceedingly rare.
2.2 Field observations
The population reported here was discovered during a field survey in the forested ravine immediately behind Nanhua Temple, located in Shaoguan City, Guangdong Province, southern China (Fig. 1). Nanhua Temple is one of the most historically significant Chan Buddhist monasteries in China, founded in 502 CE and closely associated with the Sixth Patriarch Huineng (Li, 2025). The surrounding forest has been continuously maintained as a sacred grove for over a millennium, lending the site exceptional antiquity as a cultural landscape.

Figure 1Population status and habitat conditions of Glyptostrobus pensilis at Nanhua Temple. The micro-environment of the population is characterized by severely degraded hydrological and substrate conditions: following infrastructure modifications associated with temple development, the original stream channel has ceased to flow and the surrounding ground surface has been extensively hardened (A). Canopy closure is pronounced, with the adjacent slopes dominated by subtropical evergreen broad-leaved species including Castanopsis hystrix, Camphora officinarum, and Ficus microcarpa. Advanced structural decay of the root system is visible in a representative individual (B), indicative of prolonged physiological deterioration. Canopy assessment reveals severe crown loss across the population: one individual exhibits a highly fragmented residual crown confined to scattered lateral branches, while an adjacent specimen has undergone complete mortality with total canopy absence (C).
A total of nine individual G. pensilis trees were recorded within the ravine. All were mature or senescent individuals; no seedlings, saplings, or juvenile trees of any size class were observed, indicating a complete absence of natural regeneration. Dendrochronological analysis reveals that the most ancient individual dates back to the Ming Dynasty with an estimated age of nearly 500 years (Li et al., 2004), consistent with the long history of passive protection afforded by the temple grounds.
The physiological condition of the population was severely compromised. Five of the nine individuals exhibited advanced deterioration: extensive bark exfoliation, marked crown dieback with foliage restricted to scattered apical clusters, and evidence of structural decay in major scaffold branches. At least two individuals appeared to have died entirely prior to the survey, with standing deadwood and collapsed crown architecture. Of the remaining trees, few displayed canopy conditions consistent with normal physiological function. No reproductive structures (cones or seeds) were observed on any individual during the survey period.
The population occupies a narrow valley floor position within a gorge oriented roughly west–east. The southern slope of the ravine supports large emergent trees including Toona ciliata and other tall broad-leaved species, while the northern slope is dominated by Castanopsis hystrix, Camphora officinarum, and Ficus microcarpa. The combined effect of these opposing slopes is severe light limitation at the valley floor throughout much of the day, creating a deeply shaded microenvironment that is suboptimal for G. pensilis, a species requiring relatively high light levels for successful regeneration. The canopy closure of the surrounding vegetation has progressively intensified as the sacred grove has matured in the absence of disturbance or management, further suppressing any potential for natural seedling establishment.
The population thus displays all diagnostic criteria of demographic collapse: a truncated age structure composed exclusively of senescent adults, zero recruitment, high individual mortality, and a microhabitat that is structurally unsuitable for regeneration.
2.3 Habitat characteristics
The demographic collapse of the G. pensilis micro-population at Nanhua Temple is fundamentally driven by the progressive degradation of its localized habitat (Fig. 2). While this relict species is ecologically adapted to riparian and wetland environments, the current site conditions have deviated significantly from its biological requirements. The surviving individuals are confined to a highly modified landscape, primarily clustered around the “Nine-Dragon Pool”, where natural hydrological connectivity has been severed by the construction of concrete embankments and paved roads. Such infrastructure restricts natural water level fluctuations and compromises the soil aeration essential for root health. Beyond physical habitat fragmentation, the population is subjected to significant hydro-chemical stress as evidenced by sulfate concentrations in surrounding water bodies reaching as high as 12.86 mg L−1, which, when coupled with the region's historical vulnerability to acid rain and recorded precipitation pH values as low as 3.46, directly undermines the physiological resilience of these ancient conifers (Li et al., 2004). Furthermore, intense anthropogenic interference from the temple's high-traffic religious activities has exacerbated environmental stress (Liu, 2024). Inappropriate management practices, including ground hardening and the accumulation of pollutants from incense burning, have led to severe soil compaction and atmospheric degradation. These cumulative factors have inhibited natural seedling recruitment, resulting in a functionally senescent population that persists biologically but lacks the capacity for self-replacement.
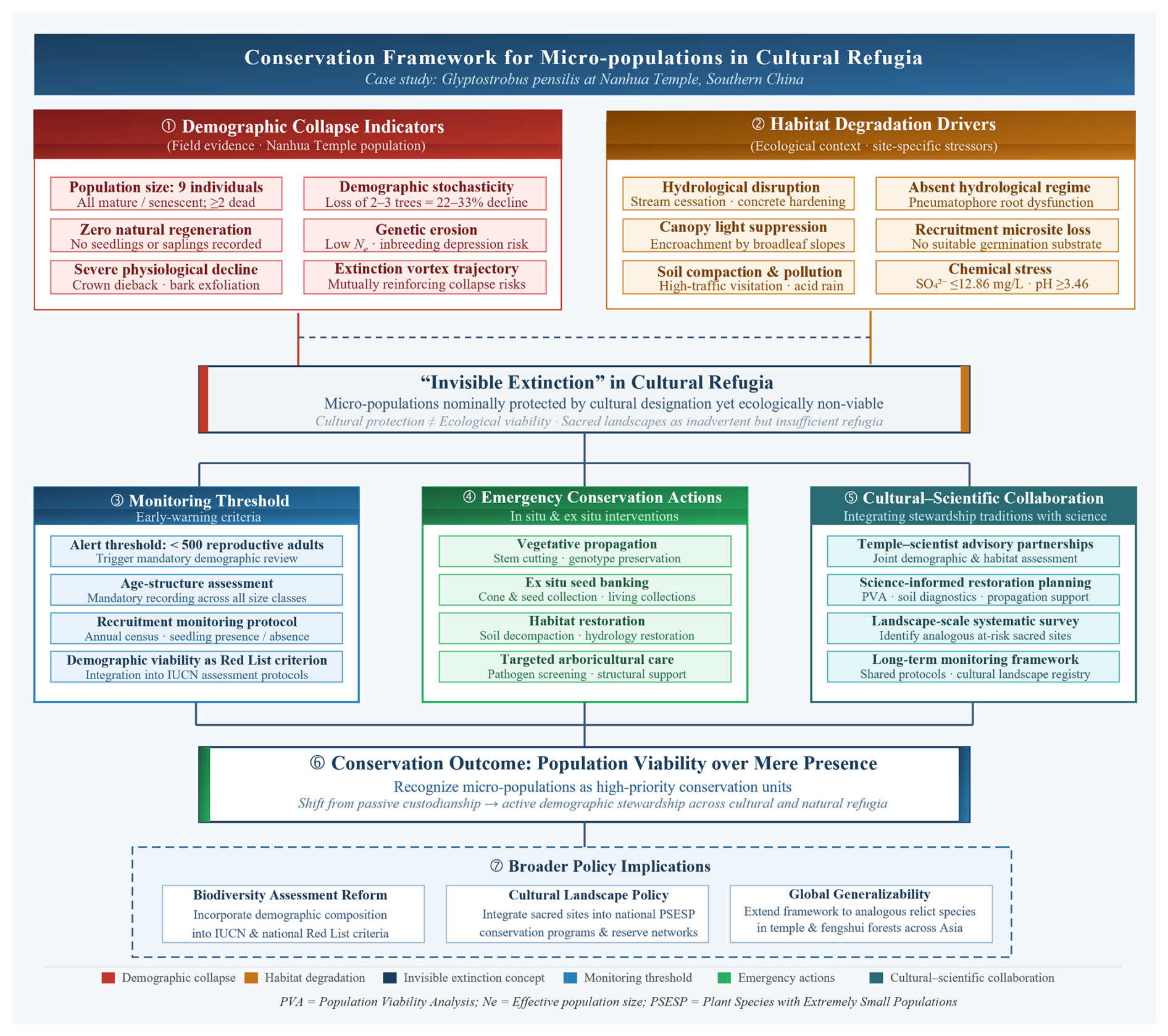
Figure 2Conceptual framework for the conservation of micro-populations in cultural refugia, illustrated by the case of Glyptostrobus pensilis at Nanhua Temple, southern China. The framework integrates field-based diagnosis with tiered conservation responses across seven functional levels. The upper tier (①②) summarizes the dual drivers of population collapse: demographic deterioration – characterized by nine senescent individuals, zero recruitment, and accelerating mortality – and progressive habitat degradation through hydrological disruption, canopy encroachment, and substrate compaction. These converging pressures underpin the central concept of “invisible extinction in cultural refugia” (③), wherein micro-populations remain nominally extant under cultural protection yet are ecologically non-viable. The middle tier outlines three complementary conservation pillars: the establishment of a demographic alert threshold (<500 reproductive individuals) to trigger mandatory management intervention (④); coordinated emergency actions spanning ex situ propagation, seed banking, habitat restoration, and targeted arboricultural support (⑤); and the formalization of cultural–scientific partnerships between temple administrators and conservation practitioners (⑥). These pillars converge toward the overarching conservation objective of prioritizing population viability over taxonomic presence (⑦), with broader policy implications for biodiversity assessment reform, integration of sacred sites into national conservation programs, and the extension of this framework to analogous relict populations across culturally managed landscapes in Asia.
3.1 Cultural protection ≠ ecological viability
The role of sacred groves, temple forests, and fengshui woodlands as de facto nature reserves has been widely documented across Asia (Sharma and Kumar, 2021; Xie et al., 2025; Huang et al., 2023; De Lacy and Shackleton, 2017). In many regions, these culturally protected landscapes constitute the last refuges for species that have been eliminated from surrounding agricultural or urban matrices (Monteagudo Churata et al., 2023; Xie et al., 2025). Their conservation value is real and should not be dismissed. However, the relationship between cultural protection and ecological viability is neither linear nor permanent, and conflating the two leads to a dangerous conservation complacency.
Historically, these sacred sites were often embedded within a more permeable and less fragmented landscape matrix. Traditional low-impact management, guided by religious taboos, unintentionally maintained a functional ecological equilibrium by allowing for natural hydrological fluctuations and seed dispersal across site boundaries. Under these pre-modern conditions, passive cultural protection was sufficient because the broader environmental processes supporting the species remained intact.
Sacred forests confer protection primarily against direct exploitation – felling, clearing, and conversion (Sullivan et al., 2024; Sharma and Kumar, 2021; De Lacy and Shackleton, 2017). They do not protect against the subtler but equally consequential processes of demographic erosion and habitat degradation. A forest can be entirely free from logging and still lose its most ecologically sensitive species through successional change (Fraver et al., 2009), but this risk is acutely amplified in contemporary settings where cultural refugia have become “islands” of senescence. The failure of modern cultural refuges stems from a critical transition: the shift from a semi-natural buffer to a highly modified, “hardened” landscape. As temple infrastructure has expanded, the original riparian micro-habitats have been replaced by concrete embankments and paved surfaces, severing the very hydrological connectivity that ancient G. pensilis individuals require for seedling recruitment. Consequently, traditional passive custodianship, while still effective at preventing logging, is now structurally incapable of counteracting the physiological stress imposed by modern environmental modification. The Nanhua Temple population of G. pensilis illustrates this precisely: the trees have survived human disturbance for centuries within the protected precinct, only to now face the slow-motion collapse of demographic non-viability and physiological senescence.
Furthermore, the conditions within many sacred groves have changed considerably from those that prevailed when culturally significant trees were first established. Infrastructure development, changes in drainage, increased visitation, and the natural development of surrounding vegetation have collectively altered the ecological context in ways that may fundamentally undermine the habitat requirements of the very species these landscapes have historically sheltered (Liu, 2024; Li, 2025). Passive protection through the simple maintenance of taboos against cutting or disturbance fails to compensate for these structural ecological changes.
3.2 Demographic and genetic risks in micro-populations
The nine surviving individuals of G. pensilis at Nanhua Temple represent not merely a small population but a population in an advanced state of demographic dysfunction. Understanding why this matters requires brief consideration of the population biology of extreme rarity.
In any population, individuals die and are replaced through reproduction and recruitment. The long-term persistence of a population depends on the balance between these rates (Lande, 1993). In the Nanhua population, this balance has broken down entirely: mortality is ongoing and accelerating, while recruitment has been absent for an indeterminate but evidently extended period. The result is a population on a deterministic trajectory toward zero, independent of any external stochastic perturbation. Even if all nine surviving trees were to persist for another century, the population would still disappear without intervention.
Beyond deterministic demography, micro-populations face heightened vulnerability to stochastic extinction (Frankham, 2005). Demographic stochasticity becomes increasingly important as population size declines: in a population of nine individuals, the random death of two or three trees in a single storm, drought, or disease event represents a loss of 20 %–33 % of the total. The probability of such catastrophic events, and the capacity of a small population to recover from them, is fundamentally different from that of a large, distributed population. The persistence of only nine individuals, including several in a moribund state, indicates that the effective population size (Ne) has fallen below the critical threshold required for long-term viability. Consequently, stochastic genetic drift likely erodes allelic diversity at a rate that far exceeds any potential replenishment through gene flow, thereby accelerating the progression of an extinction vortex (Exposito-Alonso et al., 2022; Pinto et al., 2024). While the Nanhua population has ceased natural recruitment in situ, its genetic legacy remains a critical reservoir of the species' total adaptive potential. Genetic approaches in this context are not focused on the immediate evolution of the current individuals, but rather on “genetic rescue” – the preservation of unique genotypes through vegetative propagation and germplasm banking. This ensures that even if the Nanhua stand collapses demographically, its biological information is not permanently lost and can serve as a “source” for future reintroduction efforts in more suitable riparian habitats. Inbreeding depression may further reduce the fitness of any seeds that are produced, compounding the barriers to regeneration (Li et al., 2012). The population is, in all demographic and genetic respects, functionally non-viable.
3.3 Invisible extinction in cultural refugia
We define “invisible extinction in cultural refugia” as the phenomenon where a biological population, although nominally protected by cultural or religious status, undergoes a continuous and irreversible demographic decline due to the absence of proactive ecological management – a process that remains undetected by conventional conservation monitoring systems. This situation at Nanhua Temple is not an isolated anomaly but rather an instance of a more general and under-recognized pattern. Across subtropical and tropical Asia, a disproportionate number of threatened tree species are now known only, or primarily, from populations embedded in sacred or culturally protected landscapes. A brief survey of the literature reveals analogous situations: elderly individuals of locally threatened species persisting in temple courtyards, monastery forests, or village fengshui groves, celebrated as heritage trees but slowly dying without replacement (Kendal et al., 2012; Rival, 1998).
We propose the term “invisible extinction in cultural refugia” to describe this pattern: the gradual, unmonitored disappearance of micro-populations that are nominally protected by cultural designation but are ecologically non-viable. The “invisibility” of this phenomenon has several dimensions. Several factors contribute to the overlooking of these populations in conservation strategies. First, their continued physical presence often fails to trigger Red List reassessments, as periodic evaluations frequently lack the demographic resolution necessary to capture fine-scale population declines. Second, the conventional framing of temple groves as icons of harmonious human–nature coexistence fosters a perception of security that discourages rigorous assessment of actual population viability. Third, the lack of systematic monitoring ensures that physiological deterioration proceeds without quantitative documentation, rendering local extinction a retrospective realization rather than a preventable outcome.
The global extent of this phenomenon remains unquantified but is likely substantial. Numerous culturally significant tree species across East and Southeast Asia, including various Dalbergia spp., Aquilaria spp., and rare conifers such as G. pensilis, possess populations that exhibit the functional senescence described in this study. Without a systematic effort to audit the demographic trajectories of these culturally embedded populations, conservation planning will continue to suffer from a false sense of security, leading to significant overestimations of species' long-term viability.
4.1 Establish micro-population monitoring thresholds
Contemporary conservation monitoring frameworks predominantly assess population viability using minimum viable population (MVP) theory, which has historically posited that populations numbering fewer than 50–500 individuals experience heightened extinction risks across demographic and genetic timescales (Franklin, 1980; Shaffer, 1981). While subsequent meta-analyses have suggested substantially higher median MVP estimates (Traill et al., 2007), and recent genetic studies recommend revised thresholds of 100 and 1000 individuals to mitigate short-term inbreeding depression and maintain long-term evolutionary potential, respectively (Frankham et al., 2014). While these thresholds remain valuable conceptual benchmarks, they are rarely operationalized in field monitoring or conservation management protocols, particularly for tree species in cultural landscapes.
We advocate for the formal adoption of a micro-population alert threshold, defined as fewer than 500 mature individuals at a population. This threshold is aligned with the operational definition of Plant Species with Extremely Small Populations (PSESP) in China, a conservation paradigm designed to identify populations that have fallen below the functional levels of self-sustaining viability (Yang et al., 2020). The choice of 500 individuals serves as a critical demographic “red line” because, for long-lived conifers like G. pensilis, populations below this size are highly susceptible to acute genetic drift and environmental stochasticity, leading to the “invisible extinction” observed at Nanhua Temple. This threshold is deliberately conservative and operationally straightforward, specifically designed to capture clusters such as the Nanhua G. pensilis population that have clearly fallen below the lower bounds of self-sustaining viability yet remain unflagged by existing monitoring systems. Populations failing to meet this numerical standard should automatically qualify for expedited conservation status reviews, immediate resource allocation, and strategic intervention planning, irrespective of their location within formally designated protected areas. Furthermore, institutionalizing demographic composition, particularly the presence or absence of recruitment across multiple age classes, should be institutionalized as a mandatory reporting criterion in all population monitoring surveys to complement the traditional counts of adult individuals.
4.2 Emergency conservation actions for the Nanhua population
- I.
Urgent genetic rescue and ex situ propagation. For the Nanhua G. pensilis population, intervention opportunities are rapidly diminishing. We recommend immediate genetic rescue through two complementary pathways. Given the advanced senescence of surviving individuals, vegetative propagation via stem cuttings offers the most pragmatic route to preserve extant genotypes (Wojnicka-Półtorak et al., 2022). To capture residual genetic diversity, all reproductive material including cones and seeds should be systematically banked ex situ (Exposito-Alonso et al., 2022; León-Lobos et al., 2012). Resulting progeny must be maintained in controlled environments until sufficiently robust for reintroduction to riparian zones adjacent to the original site, or retained in living collections as demographic insurance against localized extinction (Griffith et al., 2015).
- II.
Habitat restoration, individual revitalization, and systematic monitoring. In tandem with ex situ efforts, in situ habitat restoration is essential to recreate conditions conducive to seedling establishment (Grossnickle, 2018). Critical actions include targeted soil decompaction to improve aeration, drainage management to restore seasonal hydrological fluctuations, and selective thinning of competing vegetation to enhance light availability for the remaining canopy. To maximize the functional lifespan of the most compromised individuals as seed sources, we recommend implementing targeted arboricultural care including pathogen screening, structural stabilization, and the application of soil amendments (Fay, 2002). Furthermore, a long-term demographic monitoring framework must be established (Schweiger et al., 2020), utilizing annual censuses to track physiological vitality, reproductive output, and the success of any natural or assisted recruitment.
4.3 Integrate sacred-site management with conservation science
Sacred forests and temple groves have long been managed by religious communities as expressions of cultural and spiritual values (Sullivan et al., 2024; Xie et al., 2025; Frascaroli et al., 2016), and it is precisely this sustained stewardship that has allowed populations such as the G. pensilis grove at Nanhua Temple to endure into the present. The challenge, therefore, is not to replace or override existing cultural management, but to complement it with the ecological knowledge and technical capacity that passive custodianship alone cannot provide.
We call for strengthened collaboration between conservation biologists and the administrators of culturally protected landscapes harboring threatened plant populations (Doda Doffana, 2019). Such collaboration should be practical and site-specific: joint demographic assessments, shared monitoring protocols, and technically guided habitat restoration implemented in a manner consistent with the cultural character of the site. In instances where temple or monastery managers already exercise stewardship over an endangered population, scientific partners can provide the diagnostic tools including population viability analysis, soil assessment, and propagation expertise required to transform well-intentioned protection into ecologically effective conservation. Furthermore, while this study focuses on ecological diagnostics, future conservation strategies should adopt mixed-methods approaches that integrate social and anthropological perspectives to fully capture the complex human-nature interactions in temple forests (Xie and Jim, 2026).
More broadly, this case draws attention to a category of threatened population that deserves far greater systematic attention: micro-populations of endangered species persisting within culturally managed landscapes beyond the boundaries of formal nature reserves. Sites analogous to Nanhua Temple almost certainly exist across subtropical China and the wider region, their resident populations equally fragile and equally overlooked. We urge botanists, conservation practitioners, and heritage site administrators alike to survey such landscapes with demographic rigor, to recognize regeneration failure as an emergency signal rather than a background condition, and to treat the living trees in their care not merely as cultural monuments, but as the last biological representatives of populations that cannot afford further delay.
The remnant micro-population of G. pensilis at Nanhua Temple serves as a stark reminder that conservation status and conservation security are not synonymous. Nine senescent trees, sheltered for centuries by cultural reverence yet devoid of regeneration and on an irreversible demographic trajectory, expose a critical gap in how we identify, assess, and respond to extinction risk in threatened plant species. Micro-populations embedded in sacred or cultural landscapes are particularly susceptible to this gap: their cultural significance confers visibility and nominal protection while simultaneously obscuring the ecological dysfunction unfolding within them. Addressing this requires three urgent shifts in conservation practice – recognizing demographic viability, not mere presence, as the operative measure of a population's security; treating micro-populations below critical size and recruitment thresholds as conservation emergencies rather than low-priority residuals; and building genuine partnerships between conservation scientists and cultural institutions to transform passive custodianship into active ecological stewardship. Without timely intervention, many such remnant populations may disappear silently before their loss is even recognized.
No data sets were used in this article.
CX conceived the study and wrote the original draft; DL reviewed and edited the manuscript.
The contact author has declared that neither of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
The authors thank the two anonymous reviewers whose feedback greatly improved the quality and clarity of the article.
This research was supported by the National Natural Science Foundation of China (grant no. 32360417), and the Hainan Provincial Natural Science Foundation of China (grant no. 725RC789).
This paper was edited by Sonja Knapp and reviewed by two anonymous referees.
Ben, A. L.: The Taxonomy and Biogeographic History of genus-species Glyptostrobus Endlicher (Cupressaceae), B. Peabody Mus. Nat. Hi., 48, 359–426, https://doi.org/10.3374/0079-032X(2007)48[359:TTABHO]2.0.CO;2, 2007.
Crist, E.: Witnessing mass extinction: What's invisible, what's visible, what's possible, Biol. Conserv., 275, 109696, https://doi.org/10.1016/j.biocon.2022.109696, 2022.
De Lacy, P. and Shackleton, C. M.: Woody plant species richness, composition and structure in urban sacred sites, Grahamstown, South Africa, Urban Ecosyst., 20, 1169–1179, https://doi.org/10.1007/s11252-017-0669-y, 2017.
Doda Doffana, Z.: Sacred sites and ancestor veneration in Sidama, southwest Ethiopia: A socio-ecological perspective, Cogent Social Sciences, 5, 1704600, https://doi.org/10.1080/23311886.2019.1704600, 2019.
Ensslin, A. and Godefroid, S.: How the cultivation of wild plants in botanic gardens can change their genetic and phenotypic status and what this means for their conservation value, Sibbaldia: the International Journal of Botanic Garden Horticulture, 51–70, https://doi.org/10.24823/Sibbaldia.2019.267, 2019.
Exposito-Alonso, M., Booker, T. R., Czech, L., Gillespie, L., Hateley, S., Kyriazis, C. C., Lang, P. L. M., Leventhal, L., Nogues-Bravo, D., Pagowski, V., Ruffley, M., Spence, J. P., Toro Arana, S. E., Weiß, C. L., and Zess, E.: Genetic diversity loss in the anthropocene, Science, 377, 1431–1435, https://doi.org/10.1126/science.abn5642, 2022.
Fay, N.: Environmental arboriculture, tree ecology and veteran tree management, Arboricultural Journal, 26, 213–238, https://doi.org/10.1080/03071375.2002.9747336, 2002.
Frankham, R.: Genetics and extinction, Biol. Conserv., 126, 131–140, https://doi.org/10.1016/j.biocon.2005.05.002, 2005.
Frankham, R., Bradshaw, C. J. A., and Brook, B. W.: Genetics in conservation management: Revised recommendations for the 50/500 rules, Red List criteria and population viability analyses, Biol. Conserv., 170, 56–63, https://doi.org/10.1016/j.biocon.2013.12.036, 2014.
Franklin, I. R.: Evolutionary change in small populations, in: Conservation Biology: An Evolutionary-Ecological Perspective, edited by: Soulé, M. E. and Wilcox, B. A., Sinauer, Sunderland, MA, 135–149, ISBN 0878938001, 1980.
Frascaroli, F., Bhagwat, S., Guarino, R., Chiarucci, A., and Schmid, B.: Shrines in Central Italy conserve plant diversity and large trees, Ambio, 45, 468–479, https://doi.org/10.1007/s13280-015-0738-5, 2016.
Fraver, S., White, A. S., and Seymour, R. S.: Natural disturbance in an old-growth landscape of northern Maine, USA, J. Ecol., 97, 289–298, https://doi.org/10.1111/j.1365-2745.2008.01474.x, 2009.
Gilpin, M. E. and Soulé, M. E.: Minimum viable populations: Processes of extinction, in: Conservation Biology: The Science of Scarcity and Diversity, edited by: Soulé, M. E., Sinauer Associates, Sunderland, Massachusetts, USA, 19–34, ISBN 0878937943, 1986.
Griffith, M. P., Calonje, M., Meerow, A. W., Tut, F., Kramer, A. T., Hird, A., Magellan, T. M., and Husby, C. E.: Can a botanic garden cycad collection capture the genetic diversity in a wild population?, Int. J. Plant Sci., 176, 1–10, https://doi.org/10.1086/678466, 2015.
Grossnickle, S.: Seedling Establishment on a Forest Restoration Site: An Ecophysiological Perspective, Reforesta, 110–139, https://doi.org/10.21750/REFOR.6.09.62, 2018.
Hao, Z., Cheng, T., Zheng, R., Xu, H., Zhou, Y., Li, M., Lu, F., Dong, Y., Liu, X., Chen, J., and Shi, J.: The Complete Chloroplast Genome Sequence of a Relict Conifer Glyptostrobus pensilis: Comparative analysis and insights into dynamics of chloroplast genome rearrangement in Cupressophytes and Pinaceae, Plos One, 11, e0161809, https://doi.org/10.1371/journal.pone.0161809, 2016.
Huang, L., Jin, C., Pan, Y., Zhou, L., Hu, S., Guo, Y., Meng, Y., Song, K., Pang, M., Li, H., Lin, D., Xu, X., Minor, J., Coggins, C., Jim, C. Y., Yan, E., Yang, Y., Tang, Z., and Lindenmayer, D. B.: Human activities and species biological traits drive the long-term persistence of old trees in human-dominated landscapes, Nat. Plants, 9, 898–907, https://doi.org/10.1038/s41477-023-01412-1, 2023.
Kendal, D., Williams, N. S. G., and Williams, K. J. H.: A cultivated environment: Exploring the global distribution of plants in gardens, parks and streetscapes, Urban Ecosyst., 15, 637–652, https://doi.org/10.1007/s11252-011-0215-2, 2012.
Lande, R.: Risks of Population Extinction from Demographic and Environmental Stochasticity and Random Catastrophes, Am. Nat., 142, 911–927, https://doi.org/10.1086/285580, 1993.
Ledford, H.: World's largest plant survey reveals alarming extinction rate, Nature, 570, 148–149, https://doi.org/10.1038/d41586-019-01810-6, 2019.
León-Lobos, P., Way, M., Aranda, P. D., and Lima-Junior, M.: The role of ex situ seed banks in the conservation of plant diversity and in ecological restoration in Latin America, Plant Ecol. Divers., 5, 245–258, https://doi.org/10.1080/17550874.2012.713402, 2012.
Leroy, G., Carroll, E. L., Bruford, M. W., DeWoody, J. A., Strand, A., Waits, L., and Wang, J.: Next-generation metrics for monitoring genetic erosion within populations of conservation concern, Evol. Appl., 11, 1066–1083, https://doi.org/10.1111/eva.12564, 2018.
Li, F. and Xia, N.: The geographical distribution and cause of threat to Glyptostrobus pensilis (Taxodiaceae), Journal of Tropical and Subtropical Botany, 12, 13–20, 2004.
Li, R., Tan, H., Liu, Y., and Song, L.: Age and environment of Glyptostrobus pensilis in Nanhua Temple in Guangdong, Tropical Geography, 24, 321–325, 2004.
Li, W.: A Study of temple property in mountainous area of Lingnan and regional society during the Ming and Qing Dynasties: Focusing on Nanhua Temple, Journal of Sun Yat-sen University (Social Science Edition), 65, 144–155, https://doi.org/10.13471/j.cnki.jsysusse.2025.05.015, 2025.
Li, Y.-Y., Guan, S.-M., Yang, S.-Z., Luo, Y., and Chen, X.-Y.: Genetic decline and inbreeding depression in an extremely rare tree, Conserv. Genet., 13, 343–347, https://doi.org/10.1007/s10592-011-0286-x, 2012.
Liu, Y.: Sacred Resurgence: Revitalizing Buddhist Temples in Modern China, https://doi.org/10.3390/rel15050627, 2024.
Maschinski, J., Coates, D., Monks, L., Dillon, R., Barrett, S., Possley, J., Lange, J., Duquesnel, J., Goodman, J., Hermanutz, L., and Copp, C.: Rare and Threatened Plant Conservation Translocations: Lessons Learned and Future Directions, in: Ecological Restoration: Moving Forward Using Lessons Learned, edited by: Florentine, S., Gibson-Roy, P., Dixon, K. W., and Broadhurst, L., Springer International Publishing, Cham, 287–322, https://doi.org/10.1007/978-3-031-25412-3_8, 2023.
Mastretta-Yanes, A., da Silva, J. M., Grueber, C. E., Castillo-Reina, L., Köppä, V., Forester, B. R., Funk, W. C., Heuertz, M., Ishihama, F., Jordan, R., Mergeay, J., Paz-Vinas, I., Rincon-Parra, V. J., Rodriguez-Morales, M. A., Arredondo-Amezcua, L., Brahy, G., DeSaix, M., Durkee, L., Hamilton, A., Hunter, M. E., Koontz, A., Lang, I., Latorre-Cárdenas, M. C., Latty, T., Llanes-Quevedo, A., MacDonald, A. J., Mahoney, M., Miller, C., Ornelas, J. F., Ramírez-Barahona, S., Robertson, E., Russo, I.-R. M., Santiago, M. A., Shaw, R. E., Shea, G. M., Sjögren-Gulve, P., Spence, E. S., Stack, T., Suárez, S., Takenaka, A., Thurfjell, H., Turbek, S., van der Merwe, M., Visser, F., Wegier, A., Wood, G., Zarza, E., Laikre, L., and Hoban, S.: Multinational evaluation of genetic diversity indicators for the Kunming-Montreal Global Biodiversity Framework, Ecol. Lett., 27, e14461, https://doi.org/10.1111/ele.14461, 2024.
Monteagudo Churata, L., Ramos-Baldárrago, S., and Quipuscoa Silvestre, V.: Trees in urban religious sites: An analysis of their diversity in Arequipa, Peru, Urban For. Urban Gree., 86, 128022, https://doi.org/10.1016/j.ufug.2023.128022, 2023.
Nic Lughadha, E., Bachman, S. P., Leão, T. C. C., Forest, F., Halley, J. M., Moat, J., Acedo, C., Bacon, K. L., Brewer, R. F. A., Gâteblé, G., Gonçalves, S. C., Govaerts, R., Hollingsworth, P. M., Krisai-Greilhuber, I., de Lirio, E. J., Moore, P. G. P., Negrão, R., Onana, J. M., Rajaovelona, L. R., Razanajatovo, H., Reich, P. B., Richards, S. L., Rivers, M. C., Cooper, A., Iganci, J., Lewis, G. P., Smidt, E. C., Antonelli, A., Mueller, G. M., and Walker, B. E.: Extinction risk and threats to plants and fungi, Plants, People, Planet, 2, 389–408, https://doi.org/10.1002/ppp3.10146, 2020.
Nordstrom, S. W., Hufbauer, R. A., Olazcuaga, L., Durkee, L. F., and Melbourne, B. A.: How density dependence, genetic erosion and the extinction vortex impact evolutionary rescue, P. Roy. Soc. B-Biol. Sci., 290, 20231228, https://doi.org/10.1098/rspb.2023.1228, 2023.
Pinto, A. V., Hansson, B., Patramanis, I., Morales, H. E., and van Oosterhout, C.: The impact of habitat loss and population fragmentation on genomic erosion, Conserv. Genet., 25, 49–57, https://doi.org/10.1007/s10592-023-01548-9, 2024.
Rival, L.: The Social Life of Trees: Anthropological Perspectives on Tree Symbolism, Routledge, London, UK, ISBN 9781859739280, 1998.
Rønsted, N., Walsh, S. K., Clark, M., Edmonds, M., Flynn, T., Heintzman, S., Loomis, A., Lorence, D., Nagendra, U., Nyberg, B., Opgenorth, M., Weisenberger, L., Williams, A., Wolkis, D., Wood, K. R., and Keir, M.: Extinction risk of the endemic vascular flora of Kauai, Hawaii, based on IUCN assessments, Conserv. Biol., 36, e13896, https://doi.org/10.1111/cobi.13896, 2022.
Samways, M. J., Gaigher, R., Deacon, C., and Pryke, J. S.: Heterogeneous conservation corridors of remnant vegetation protect biodiversity in South African timber mosaics, Biodivers. Conserv., 34, 357–379, https://doi.org/10.1007/s10531-024-02993-9, 2025.
Schweiger, A. H., Irl, S. D. H., Svenning, J.-C., and Higgins, S. I.: Dynamic management needs for long-lived, sporadically recruiting plant species in human-dominated landscapes, Plants, People, Planet, 2, 186–200, https://doi.org/10.1002/ppp3.10096, 2020.
Shaffer, M. L.: Minimum Population Sizes for Species Conservation, BioScience, 31, 131–134, https://doi.org/10.2307/1308256, 1981.
Sharma, S. and Kumar, R.: Sacred groves of India: repositories of a rich heritage and tools for biodiversity conservation, J. Forestry Res., 32, 899–916, https://doi.org/10.1007/s11676-020-01183-x, 2021.
Sullivan, M. K., Browne, L., Penagos Zuluaga, J. C., Liu, J., Surendra, A., and Estrada-Villegas, S.: Sacred forest biodiversity conservation: A meta-analysis, Conserv. Sci. Pract., 6, e13055, https://doi.org/10.1111/csp2.13055, 2024.
Tang, C. Q., Yang, Y., Momohara, A., Wang, H.-C., Luu, H. T., Li, S., Song, K., Qian, S., LePage, B., Dong, Y.-F., Han, P.-B., Ohsawa, M., Le, B. T., Tran, H. D., Dang, M. T., Peng, M.-C., and Wang, C.-Y.: Forest characteristics and population structure of Glyptostrobus pensilis, a globally endangered relict species of southeastern China, Plant Diversity, 41, 237–249, https://doi.org/10.1016/j.pld.2019.06.007, 2019.
Thomas, P., Yang, Y., Farjon, A., Nguyen, D., and Liao, W.: Glyptostrobus pensilis (amended version of 2011 assessment), The IUCN Red List of Threatened Species 2020, T32312A177795446, https://doi.org/10.2305/IUCN.UK.2020-3.RLTS.T32312A177795446.en, 2020.
Traill, L. W., Bradshaw, C. J. A., and Brook, B. W.: Minimum viable population size: A meta-analysis of 30 years of published estimates, Biol. Conserv., 139, 159–166, https://doi.org/10.1016/j.biocon.2007.06.011, 2007.
Wojnicka-Półtorak, A., Celiński, K., and Chudzińska, E.: Genetic resources of the oldest trees of Pinus sylvestris L. from the last natural forest in Europe, Dendrobiology, 89, 27–34, https://doi.org/10.12657/denbio.089.003, 2022.
Xie, C. and Jim, C. Y.: Bridging the gap between cultural reverence and scientific care for sacred heritage trees, Urban Ecosyst., 29, 118, https://doi.org/10.1007/s11252-026-01982-w, 2026.
Xie, C., Lyu, Y., and Jim, C. Y.: Sacred groves occluded in the urban matrix: Biodiversity and conservation potential of Hangzhou's Buddhist temples, China, Reg. Environ. Change, 25, 113, https://doi.org/10.1007/s10113-025-02450-5, 2025.
Yang, J., Cai, L., Liu, D., Chen, G., Gratzfeld, J., and Sun, W.: China's conservation program on plant species with extremely small populations (PSESP): Progress and perspectives, Biol. Conserv., 244, 108535, https://doi.org/10.1016/j.biocon.2020.108535, 2020.
Yang, X., Liang, Y., Wang, X., Deng, B., Lu, Z., Tong, D., and Liu, S.: Phenotypic diversity of wild Glyptostrobus pensilis in Guangxi and its relationship with geographical-climatic factors, Journal of Plant Resources and Environment, 33, 61–70, https://doi.org/10.3969/j.issn.1674-7895.2024.04.07, 2024.
Yang, Y., Tan, C., and Yang, Z.: Conservation of gymnosperms in China: Perspectives from the List of National Key Protected Wild Plants, Biodiversity Science, 29, 1591–1598, https://doi.org/10.17520/biods.2021403, 2021.
Ye, X., Zhang, M., Yang, Q., Ye, L., Liu, Y., Zhang, G., Chen, S., Lai, W., Wen, G., Zheng, S., Ruan, S., Zhang, T., and Liu, B.: Prediction of suitable distribution of a critically endangered plant Glyptostrobus pensilis, Forests, 13, 257, https://doi.org/10.3390/f13020257, 2022.
Zheng, Z., Ma, T., Roberts, P., Li, Z., Yue, Y., Peng, H., Huang, K., Han, Z., Wan, Q., Zhang, Y., Zhang, X., Zheng, Y., and Saito, Y.: Anthropogenic impacts on Late Holocene land-cover change and floristic biodiversity loss in tropical southeastern Asia, P. Natl. Acad. Sci. USA, 118, e2022210118, https://doi.org/10.1073/pnas.2022210118, 2021.
Zhou, L., Huang, L., Jin, C., Hu, S., Long, Y., Qian, S., Shang, K., Song, K., Song, Y.-B., Lin, D., Zhao, L., Momohara, A., and Yang, Y.: Fengshui forests as a conservation paradigm of the golden larch in China, Forest Ecol. Manag., 520, 120358, https://doi.org/10.1016/j.foreco.2022.120358, 2022.