the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 28 Aug 2019
| 28 Aug 2019
Impact of land-use change in mountain semi-dry meadows on plants, litter decomposition and earthworms
Ines Jernej
Andreas Bohner
Ronnie Walcher
Raja Imran Hussain
Arne Arnberger
Johann G. Zaller
Thomas Frank
Traditionally managed mountain grasslands are biodiversity hotspots in central Europe. However, socio-economic trends in agriculture during the last decades have changed farming practices, leaving steep and remote sites abandoned. Especially the abandonment of meadows is well known to directly affect plant and insect diversity. However, not much is known about the effects on soil processes and soil biota. To assess this, we studied four extensively managed (mown once a year, no fertilization) and four abandoned (no mowing, no fertilization) semi-dry meadows in a mountain region in Austria. Plant species richness, plant cover, plant traits, plant biomass, litter decomposition (tea bag index), and earthworm species richness and density were assessed. Additionally, soil temperature, moisture and electrical conductivity were measured. Results showed that managed meadows contained more plant species than abandoned meadows (118 vs. 93 species, respectively). We also observed different plant species assemblages between the two management types. In managed meadows, hemirosette and ruderal plant species were more abundant, while more plant species without rosettes and a higher plant necromass were found in abandoned meadows. Additionally, decomposition rate was higher in abandoned meadows. There was a trend towards higher earthworm densities in managed meadows, but there was no difference in earthworm species richness. We conclude that meadow management has effects on both aboveground vegetation and belowground biota and processes. Both abandoned and extensively managed meadows were important to sustain overall biodiversity and ecosystem functioning in the study region.
- Article
(635 KB) - Full-text XML
-
Supplement
(90 KB) - BibTeX
- EndNote





Mountain areas in central Europe consist of a high variety of natural grasslands, managed meadows and pastures, and woody areas (Schmitzberger et al., 2005). Most of the grasslands are traditionally managed and related to high species richness of, for example, plants (Schmitzberger et al., 2005), bumblebees (Walcher et al., 2017) and hoverflies (Hussain et al., 2018), thus forming biodiversity hotspots. Among these grasslands, especially semi-dry, nutrient-poor meadows represent particularly species-rich plant agroecosystems inhabiting many rare and protected plant and animal species (Bohner et al., 2003; Wilson et al., 2012). To protect these species-rich grasslands, regular extensive management is an important measure for nature conservation (Bohner et al., 2003; Lundber et al., 2017; Moog et al., 2002). However, socio-economic trends have led to a decline of traditional farming practices (Hejcman et al., 2013). Furthermore, small side-job farms are being replaced by more intensively managed larger farms (Lieskovský et al., 2015; Marini et al., 2011). Hence, two contrasting patterns can be observed: intensification (increased fertilization and mowing frequency) of easily accessible high-yielding areas on the one hand and abandonment in areas that are difficult to manage with agricultural machinery on the other hand (MacDonald et al., 2000; Niedrist et al., 2009; Tasser and Tappeiner, 2002). In this context, steep and remote mountain meadows have been increasingly abandoned (Niedrist et al., 2009) resulting in a reduction of these semi-natural grasslands within Europe (Baur et al., 2006) and a loss of these biodiversity hotspots.
Land-use changes have been shown to be the most important driving force for changes in the vegetation (Tasser and Tappeiner, 2002). These changes affect plant functional diversity and species composition (Bohner et al., 2003; Maurer, 2005; Niedrist et al., 2009; Rolecek et al., 2014; Tasser and Tappeiner, 2002) as well as the structure of plant biomass, as abandonment increases litter accumulation (Kelemen et al., 2014). Litter decomposition is a key process in ecosystems that drives nutrient cycling. Decomposition depends on changes in plant species composition, the functional characteristics of plants (Hättenschwiler et al., 2005; Meier and Bowman, 2008), the activity of soil biota such as earthworms, and abiotic factors such as temperature and moisture (Lavelle et al., 1998).
Generally, effects of grassland management on plant communities are well studied, but little is known about management impacts on soil organisms and processes and their interaction with vegetation, especially considering mountain grasslands. In many grasslands, earthworms account for most of the biomass of soil-dwelling fauna (Lavelle and Spain, 2001). They are considered as ecosystem engineers, playing an important role in soil formation and nutrient cycle (Blouin et al., 2013; Lavelle, 1997). Earthworms are known to influence the density, diversity, structure and activity of microorganisms, and soil meso- and macrofauna (Wurst et al., 2018). Earthworms have been shown to react to changes in plant diversity and functional groups (Piotrowska et al., 2013; Spehn et al., 2000; Zaller and Arnone III, 1999b), changes which have also been observed after abandonment of meadow management (e.g. changes in root biomass, nutrient supply and primary production). Consequently, we hypothesized for the current study that differences in meadow management may alter soil biota and processes via changes in plant communities (Hooper et al., 2005).
We expected that (i) mowing once a year will increase plant species richness, alter vegetation structure and reduce plant necromass in managed meadows compared to abandoned sites and that (ii) these changes in plant communities and structure will stimulate earthworm abundance and increase litter decomposition.
2.1 Study sites
The study was conducted in the central Ennstal in the Long-Term Socio-economic and Ecosystem Research (LTSER) region Eisenwurzen (federal state of Styria, Austria) across eight study sites in the municipalities of St. Gallen (47∘41′ N, 14∘37′ E), Pürgg (47∘31′ N, 14∘03′ E) and Stainach (47∘32′ N, 14∘06′ E) ranging from 660 to 790 m above sea level. All sites are located in temperate sub-oceanic climate with a long-term mean annual air temperature of 6.9 ∘C and a mean annual precipitation of 1087 mm (ZAMG, 2018). Four extensively managed and four abandoned meadows were studied in June and August 2016. Study meadows were in close distance to each other with similar site conditions such as meadow size, exposition, soil type and climate (Table S1 in the Supplement). Each managed meadow was mown once a year without any fertilizer input for at least 20 years. They are assigned to the grassland type Bromion erecti (semi-dry grassland; Mucina et al., 1993), with the grass species Bromus erectus Huds., the herb Astrantia major major L. and the grass Festuca rubra rubra L. as the dominant plant species. The abandoned meadows were not mown for on average 30 years and dominated by the tall grass Brachypodium pinnatum (L.) P. Beauv. Before abandonment, grasslands were extensively mown but not grazed. All information about management was obtained from interviews with land owners.
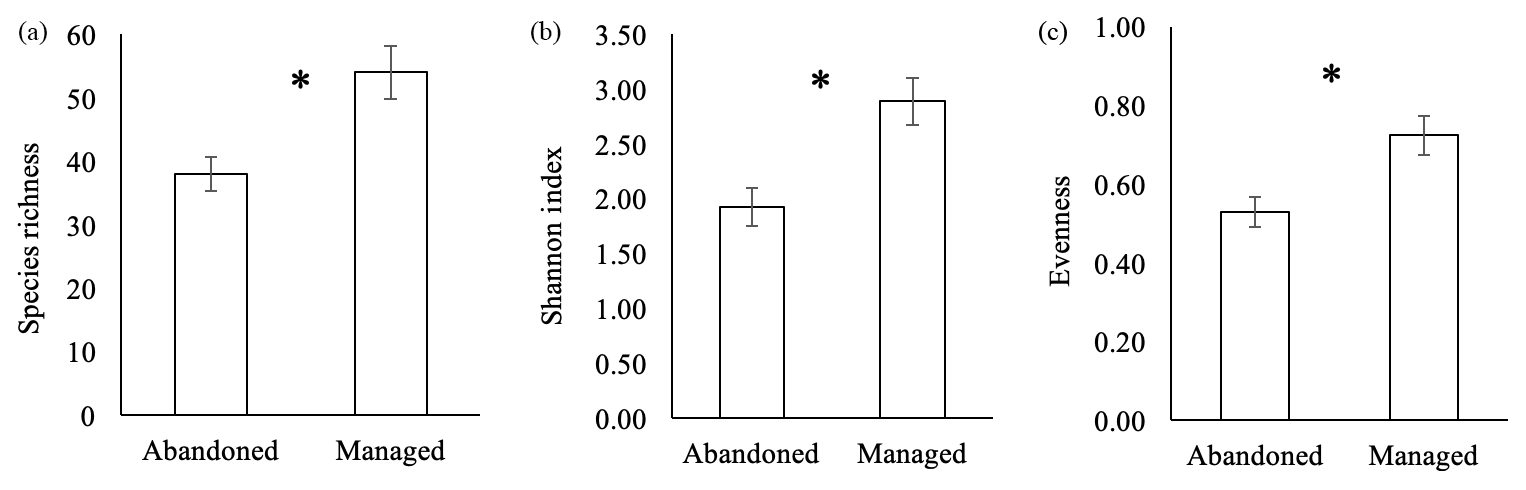
Figure 1Plant species richness (a), Shannon index (b) and evenness (c) in abandoned and managed meadows. * denotes significant difference between management types (p<0.05). Means ± SD.
2.2 Measurements
2.2.1 Vegetation
For vegetation measurements four 1 m × 1 m study plots (distance 5 m) were randomly selected on each meadow. Vascular plant species within the plots were identified (Fischer et al., 2008) and plant cover estimated using a modified Braun-Blanquet (1964) scale containing three subdivisions per cover class (Bohner et al., 2012). Plant species frequency (rooted frequency) was counted per plot.
Plant biomass was assessed in June by cutting 50 cm × 50 cm subplots within the vegetation plots (4 cm above ground) and plant material separated into necromass and living biomass. In the laboratory, living biomass was further separated in the functional groups grasses (Poaceae, Juncaceae and Cyperaceae), legumes (Fabaceae) and non-legume herbs. Necromass and functional groups were dried at 60 ∘C for 72 h and weighed.
Plant species were assigned into strategy types competitors, stress tolerators and ruderals according to Grime (1977). The management influence on plant architecture categories rosette, hemirosette and erosulate (no rosette) was assessed according to Klotz et al. (2002).
2.2.2 Litter decomposition
Following Keuskamp et al. (2013) the tea bag approach with two types of commercial tea bags (rooibos tea – Lipton, EAN: 87 22700 18843 8 and green tea – Lipton, EAN: 87 22700 05552 5) was used to determine microbial and mesofaunal litter decomposition. First, tea bags were dried for 1 h at 70 ∘C and weighed. In June, 10 tea bag pairs of rooibos and green tea were buried in 8 cm depth in a circle of 2 m radius within a representative area in each meadow considering exposition, slope, shading by trees. Second, after 68–71 days, tea bags were removed, cleaned from attached soil particles with a toothbrush, dried for 48 h at 70 ∘C and weighed. Third, according to the respective protocol (http://www.teatime4science.org/method/stepwise-protocol/, last access: 10 May 2019), decomposition rate k (rapidly decomposed plant material with easily degradable compounds) and litter stabilization factor S (labile fraction, which stabilize and become recalcitrant during decomposition) were calculated. Thirteen pairs of tea bags in managed and 4 pairs in abandoned meadows could not be found in August or were defective and not considered.
2.2.3 Earthworm sampling and soil properties
In each meadow, five sampling plots were randomly chosen at equal distance of 5 m. Next, a 25 cm × 25 cm × 25 cm (length × width × depth) soil monolith was excavated with a spade. The excavated soil was carefully searched for earthworms on a plastic foil, and each earthworm found was preserved in 4 % formol.
In the laboratory, each sample was first washed under distilled water and then separated into juvenile and adult earthworms. Juvenile earthworms without a clitellum could not be identified at the species level but were sorted into ecological groups endogeic, anecic and epigeic (Bouché, 1977), counted and weighed. Adult earthworms with a clitellum were identified after Brohmer (1984) and Christian and Zicsi (1999), also allocated to the ecological groups, counted and weighed.
Soil temperature, soil moisture and electrical conductivity were measured in parallel with each earthworm sampling using the time domain reflectometry (TRIME®-PICO 64/32, HD2, IMKO Micromodultechnik GmbH, Ettlingen, Germany). Soil pH was determined using a glass electrode on mixed soil samples (Karrer, 2015).
2.2.4 Statistical analyses
Effects of the management type (managed vs. abandoned) were examined using generalized linear models (GLMs). A Poisson distribution was used for plant and earthworm species richness, and a Gaussian distribution was used for Shannon index, evenness, total plant biomass, necromass, legumes, herbs, grasses, plant cover, plant strategy types, erosulate, hemirosette and rosette plants, decomposition rate and stabilization factor, earthworm density, endogeic, anecic and epigeic earthworms, soil electrical conductivity, pH, soil moisture, and temperature. All statistical analyses were performed in R version 3.3.1 (R Core Team, 2016) using an α<0.05 to be considered as statistically significant and to be considered as marginally significant. To assess differences in species assemblages of plants and earthworms between abandoned and managed meadows, a principal coordinate analysis (PCoA) was conducted based on a resemblance matrix of Bray–Curtis similarity measures. PERMANOVA was computed to test for significant differences in species assemblages between abandoned and managed meadows. Residuals were permuted 9999 times under a reduced model. PCoA and PERMANOVA were performed in the Primer version 6.1.13 with PERMANOVA+ (PRIMER-E Ltd., Plymouth, UK). Potential interrelations between the tested parameters were analysed by Pearson correlations for normally distributed variables and Spearman correlations for not normally distributed variables (plant cover). Normality of distributions was checked using a Shapiro test.
In total, 93 plant species were recorded in abandoned and 118 plant species in managed meadows. Plant species richness, Shannon index and evenness were significantly higher in managed compared to abandoned meadows (Fig. 1a–c).
Table 1Plant species cover and frequency in abandoned and managed meadows. Species are ordered by descending cover. Values are means.
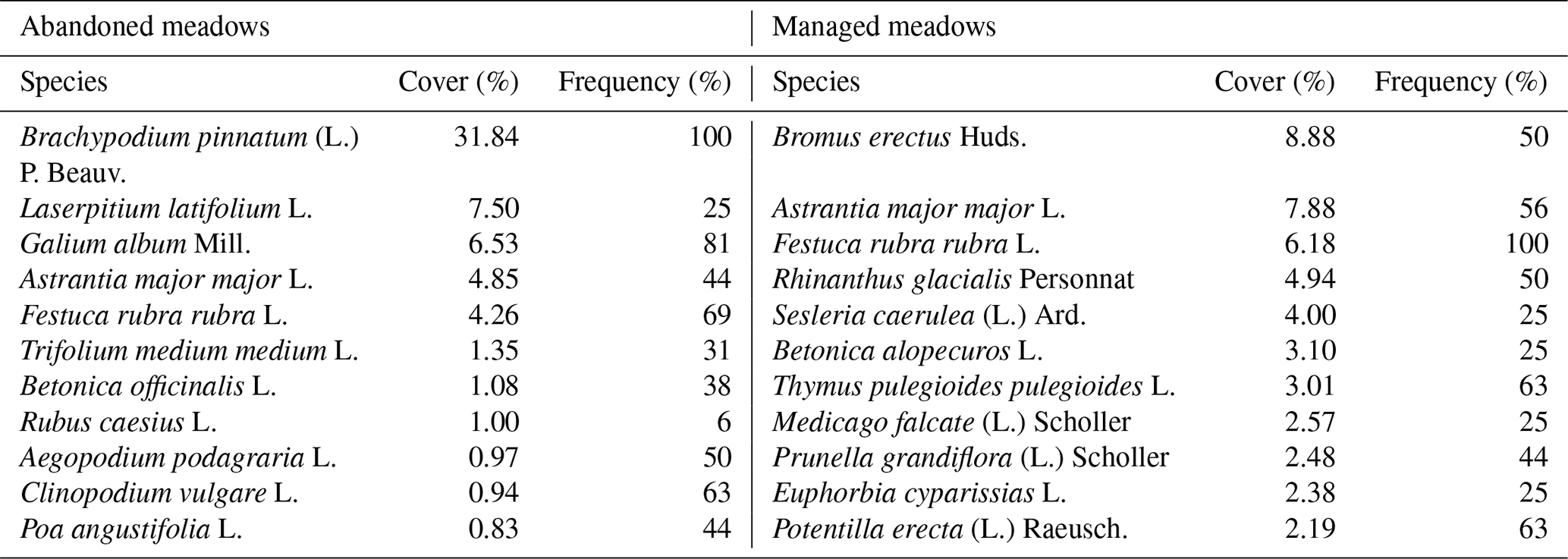
The dominant species in abandoned meadows was the grass B. pinnatum, which occurred in all sampling plots with a mean cover of 31.8 %, followed by the herb Laserpitium latifolium L. (mean cover 7.5 %; Table 1). In contrast, the cover of individual species in managed meadows was more equal. B erectus (grass), which was present in half of the plots with a mean cover of 8.9 %, was the predominant species in managed meadows, followed by the herb A. major major and the grass F. rubra rubra with a mean cover of 7.9 % and 6.2 %, respectively. Plant cover was similar in abandoned and managed sites (GLM, p=0.260).
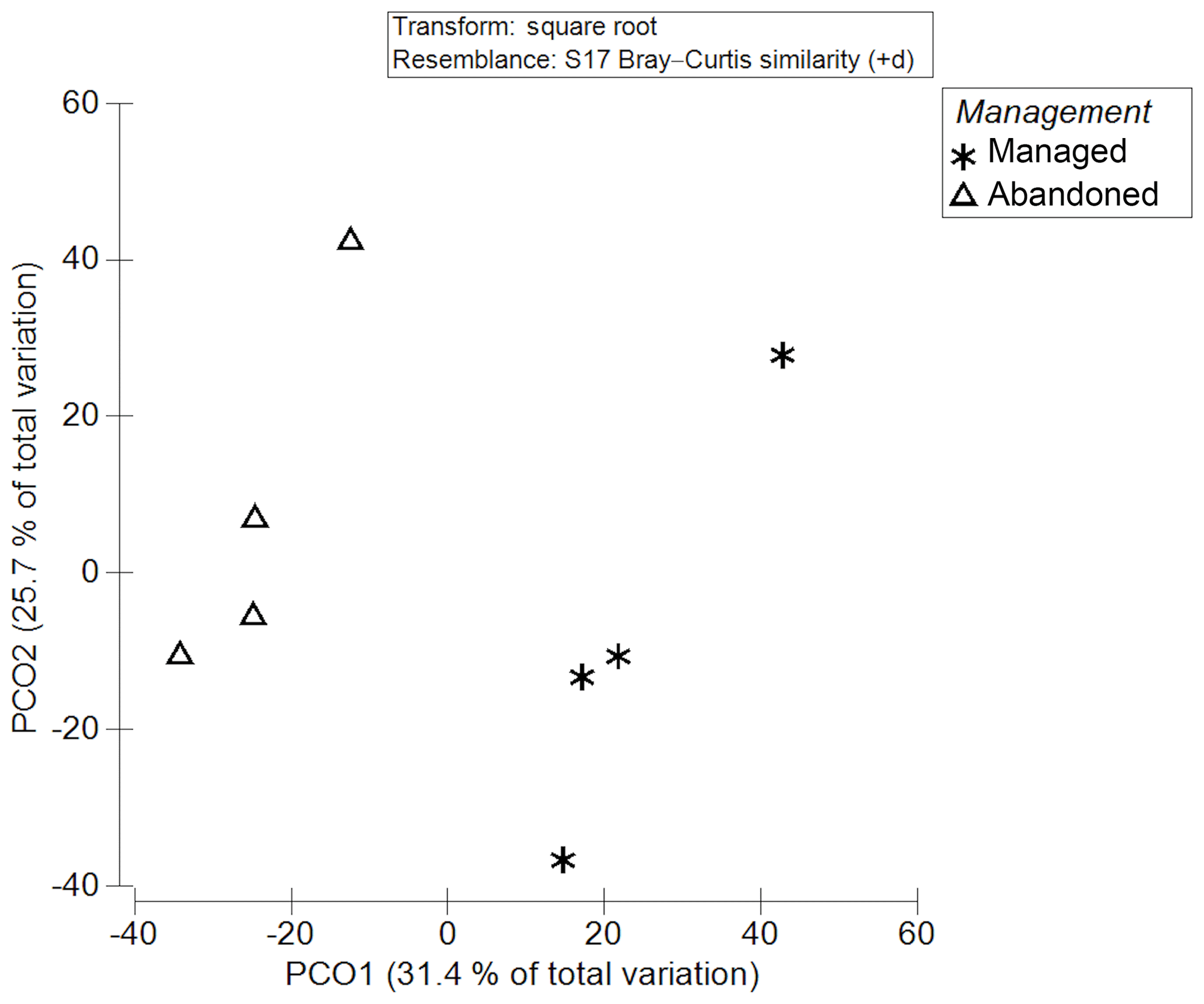
Figure 2Principal coordinate analysis (PCO = PCoA) of plant species assemblages between managed and abandoned meadows.
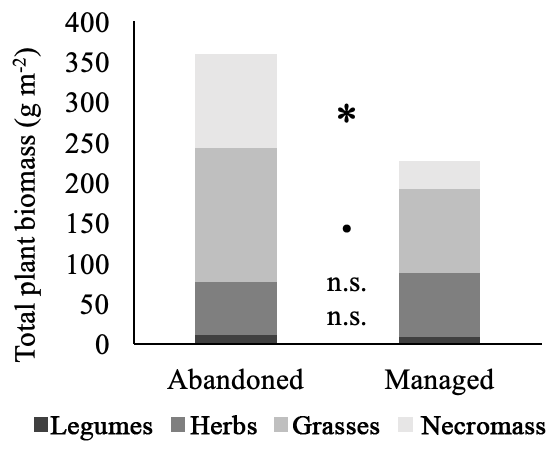
Figure 3Structure of total plant biomass in abandoned and managed meadows. * denotes significant difference between management types (p<0.05), • marginal difference (p<0.1), n.s. no significant difference. Values are means.
PCoA (Fig. 2) revealed a clear and significant separation of species assemblages between abandoned and managed meadows (PERMANOVA, p=0.028).
Total plant biomass was significantly higher in abandoned compared to managed meadows (GLM, p=0.045). Furthermore, the structure of functional groups in the plant assemblage was influenced by management (Fig. 3). Abandoned meadows had significantly higher necromass and marginally higher grass biomass. Herb and legume biomass showed no management-induced differences.
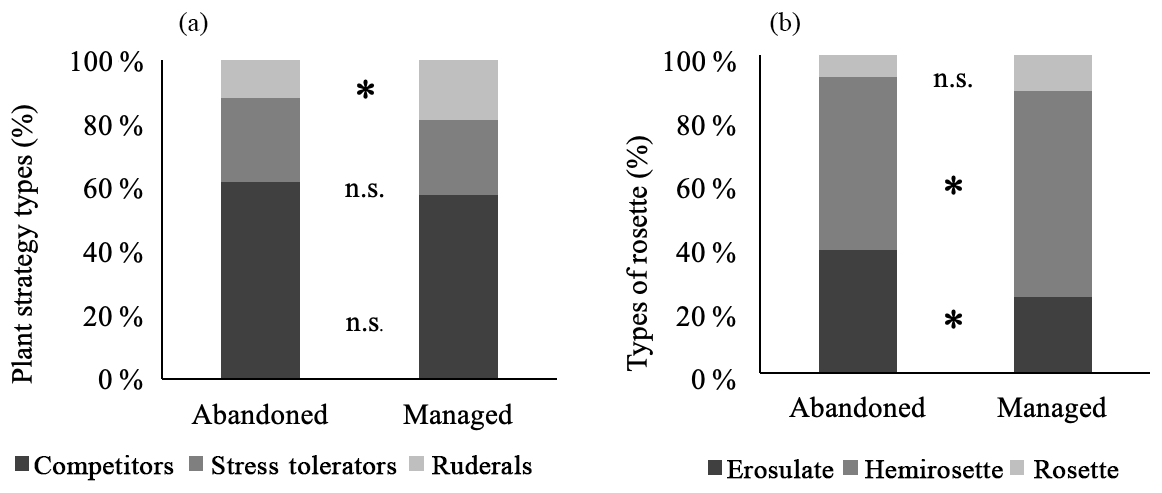
Figure 4Proportion of plant strategy types (a) and proportion of types of rosettes (b) in abandoned and managed meadows. * denotes significant difference between management types (p<0.05), n.s. no significant difference. Values are means.
Among the plant strategy types, ruderal plants had a significantly higher proportion in managed meadows. Proportions of competitors and stress tolerators were similar between management types (Fig. 4a). In abandoned meadows, significantly more erosulate growing and less hemirosette plant species were found. The predominant character states of rosettes in both management types was hemirosette (Fig. 4b). A proportion of rosette plant species showed no significant difference.
Decomposition rate k was significantly higher in abandoned meadows compared to managed meadows (Fig. 5a). For stabilization factor S, no significant difference was found (Fig. 5b).
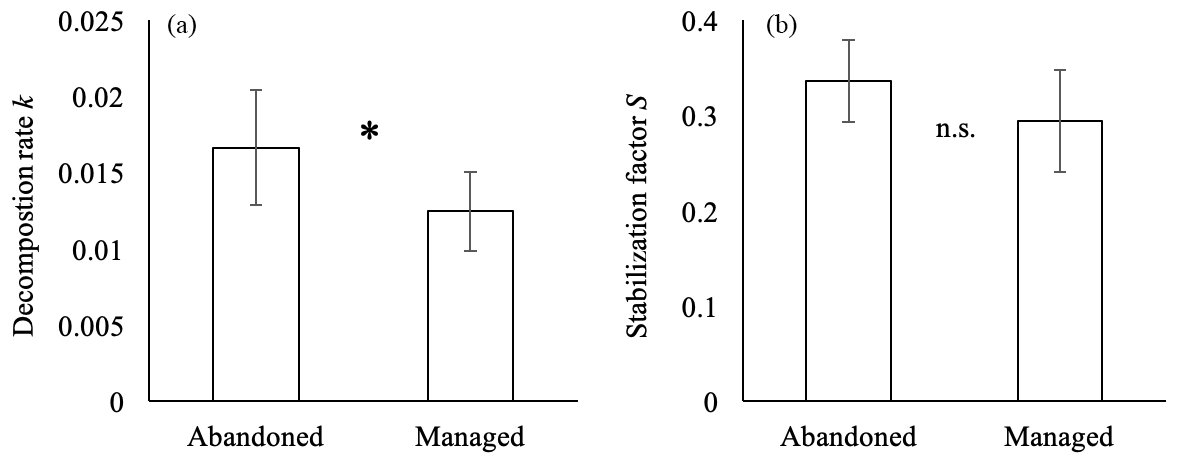
Figure 5Decomposition rate k (a) and stabilization factor S (b) measured with tea bag index in abandoned and managed meadows. * denotes significant difference between management types (p<0.05), n.s. no significant difference. Values are means ± SD.
In total, seven earthworm species were found in all meadows (Table 2). The endogeic species Aporrectodea rosea Sav. and Octolasion lacteum Oerl. were most abundant across management types, followed by the endogeic Aporrectodea caliginosa Sav. in abandoned meadows and epigeic Lumbricus rubellus Hoffm. in managed meadows. Only one species belonged to the ecological group anecic, with two recorded individuals in managed sites. Ecological groups (anecic and epigeic) did not significantly differ between meadow types. Neither species richness (GLM, p=0.866), nor earthworm assemblages were affected by management (PERMANOVA, p=0.642). Total earthworm density and endogeic earthworm density were marginally higher in managed compared to abandoned meadows (Fig. 6a, b).
Table 2Adult earthworm species found in abandoned and managed meadows with mean density of individuals per species in June and August. Values are means ± SD. Results do not include juvenile species with unclear identification.
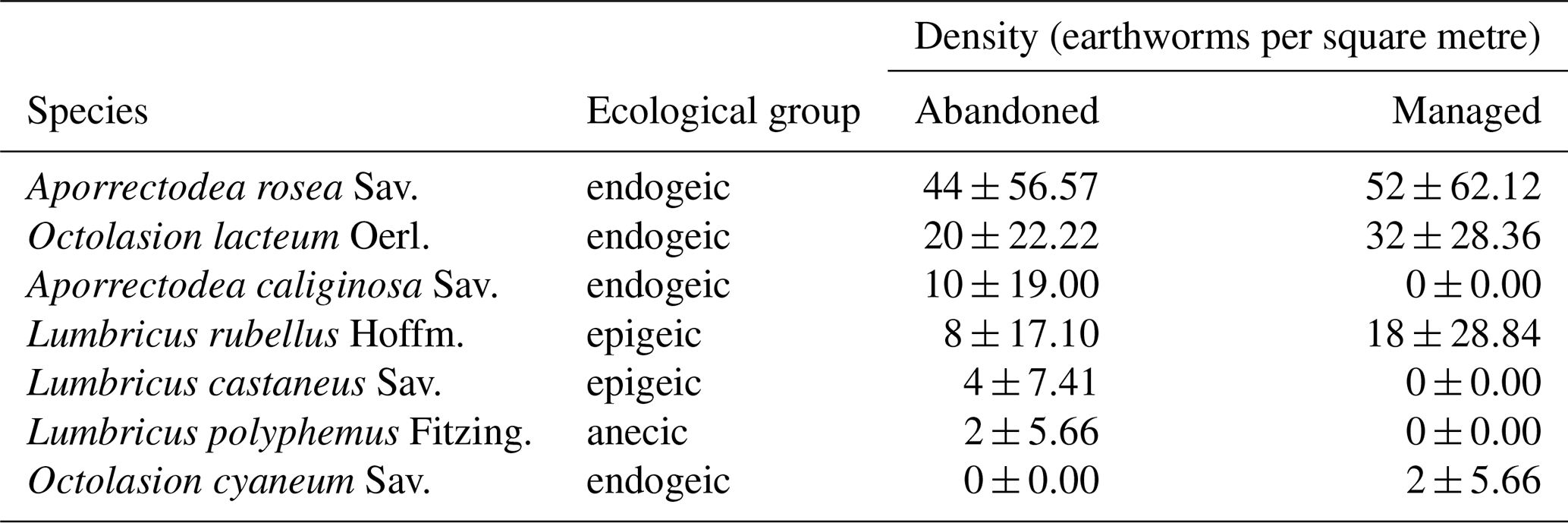
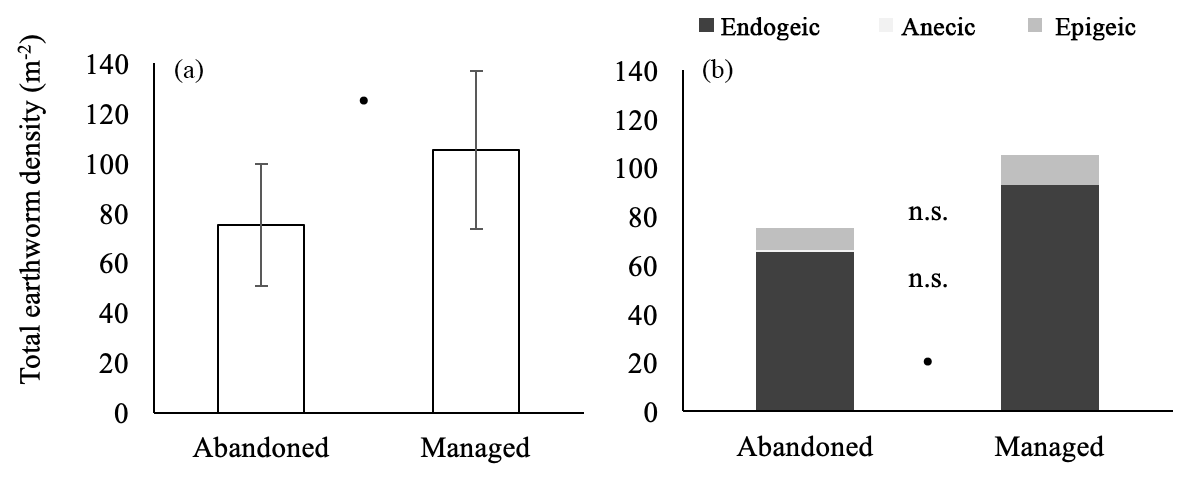
Figure 6Total earthworm density (a) and density of ecological groups (b) in abandoned and managed meadows. Anecic earthworms are shown between endogeics and epigeics but hardly depicted. • denotes marginally significant difference between management types (p<0.1), n.s. no significant difference. Values are means ± SD.
Soil temperature, soil moisture, electrical conductivity (EC) and pH did not significantly differ between management types (GLM, psoil temperature=0.858, psoil moisture=0.742, pEC=0.276, ppH=0.563).
Earthworm density showed no significant correlations with other tested parameters (Table 3). Plant species richness decreased with increasing necromass (p=0.019), decomposition rate increased with increasing necromass (p=0.004) and increasing grass biomass (p=0.014). Decomposition rate was negatively correlated with plant species richness (p=0.007). All other correlations between the measured parameters were not statistically significant.
Table 3Correlations between earthworm density, vegetation parameters and litter decomposition. Pearson and Spearman (only for plant cover) correlation coefficients. Missing results: no correlation was calculated.
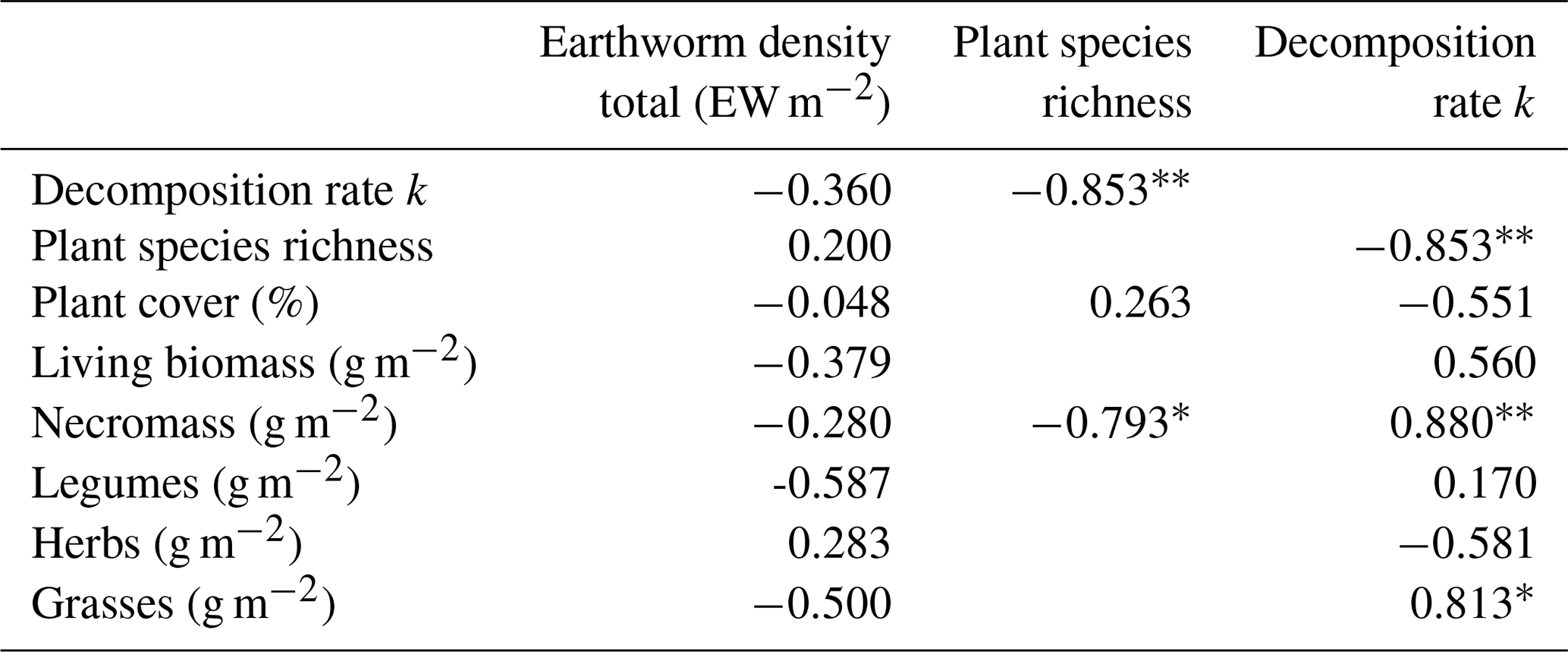
Symbols after values denote significant correlations: * p<0.05; p<0.01.
This is one of few studies considering effects of mountain meadow management on interactions between above- and belowground ecosystem biota and processes. An average of 54 plant species per square metre in extensively managed and 38 species per square metre in abandoned meadows is representative of semi-dry grasslands of the central Ennstal (Bohner et al., 2003). Likewise, the Shannon index and evenness were higher in managed meadows. This confirms other findings of decreased vascular plant species numbers due to abandonment (Fontana et al., 2014; Jacquemyn et al., 2003; Niedrist et al., 2009; Tasser and Tappeiner, 2002; Wehn et al., 2017). Disturbances in managed meadows through mowing or driving with farm machinery reduce the competitive ability of dominant plant species and create gaps for the establishment of many other species (Jacquemyn et al., 2003). The higher light availability in managed meadows enables many meso- and xerophilic, light-demanding species to co-exist, resulting in a higher species richness (Bohner et al., 2003).
Pavlů et al. (2011) argue that the principal effect of abandonment on plant species composition is based on a change in the abundance of some dominant species. In abandoned meadows, some species become dominant, forcing other species to decline (Niedrist et al., 2009). This shift of species can also be seen in abandoned meadows in the present work with a decline of the grass B. erectus (cover of 8.9 % in managed and 0.1 % in abandoned meadows) and an increase of the grass B. pinnatum (cover of 1.0 % in managed and 31.8 % in abandoned meadows). Similarly, Köhler et al. (2005) observed a negative effect of abandonment on the tufts of B. erectus, whereas B. pinnatum increased with ongoing abandonment due to benefits via more rhizomatous growth. Bobbink and Willems (1987) related the dominance of B. pinnatum to its tall growth form and aboveground phytomass. Furthermore, B. pinnatum is supposed to restrict the growth of most other plant species, resulting in a decrease of plant species diversity (Bobbink et al., 1987; Bobbink and Willems, 1987).
We also observed an accumulation of necromass in abandoned meadows and accordingly higher total plant biomass in these sites. This has also been observed by others (Kelemen et al., 2014) and has been shown to have negative effects on plant species richness, e.g. due to reduced solar irradiation on the surface, hindering germination and establishment of many plant species (Kelemen et al., 2013). Also, tall grasses are superior competitors in meadows and can suppress the biomass of most herbs (Del-Val and Crawley, 2005). The germination of grasses is less hindered by litter (Xiong and Nilsson, 1999), which makes them more resistant to cessation of management.
The decline of ruderals during succession is in line with findings by Prévosto et al. (2011), who related the shift in strategy types to the lack of disturbances after abandonment. Ruderal plants have a rapid growth rate and a high annual seed production, allowing them to establish in frequently disturbed areas. Disturbances cause a shift in the structure of the rosette types. In the present study, erosulate species increased and hemirosette species decreased during succession, suggesting that rosette plants are more sensitive to abandonment. Prévosto et al. (2011) and Weiss and Jeltsch (2015) obtained similar results. Due to less light availability in the lower canopy layers, plant types with leaves near the ground are discriminated by erosulate plants (Pavlů et al., 2011; Peco et al., 2012).
While effects of grassland management on vegetation diversity and structure have frequently been studied, little is known on potential impacts on belowground fauna and processes. We found a higher decomposition rate in abandoned than in managed meadows, indicating that a thicker layer of necromass in abandoned sites may have improved the microenvironmental conditions for decomposition (Facelli and Pickett, 1991). Similarly, Liu et al. (2009) and Novara et al. (2015) found that increasing litter quantity influences soil carbon and nitrogen cycles. Meier and Bowman (2008) showed that the composition and diversity of chemical compounds within plant litter mixtures have important effects on decomposition. However, the chemical qualities of plant litter were not considered in the present study.
Additionally, Scherer-Lorenzen (2008) observed that plant species diversity has rather weak effects on decomposition, while decomposition was found to increase with higher plant functional diversity. For example, legume abundance enhances decomposition via effects on litter quality and on the decomposition microenvironment. This is also in line with Milcu et al. (2008), who highlighted legumes as very important in grassland decomposition processes. In the present study, we found a negative impact of plant species richness on decomposition rate, while the positive effect between legume biomass and decomposition could not be observed. However, in the present results a higher biomass of grasses seemed to increase the decomposition rate. Groffman et al. (1996), who investigated the effects of grass species on microbial biomass, suggested that different grass species may influence the microbial activity, but to a lesser extent than soil type, which is considered a more important controller of microbial biomass and activity. Clearly, these comparisons should be made with caution as different approaches to determine litter decomposition have been used and different ecosystems were investigated.
We found a total of seven earthworm species in the studied meadows, which is a similar species richness than reported from grasslands of lower altitudes in Europe (Cluzeau et al., 2012; Zaller and Arnone III, 1999b). Based on a long-term field experiment, Pop (1997) concluded that 2–4 endogeic, 0–2 anecic and 1–2 epigeic species are to be expected in mountain woody and grassland ecosystems, which was also observed in the present study. In the current study, earthworm species richness and species assemblages remained unaffected by abandonment even if abandonment lasted for 20–40 years. This indicated negligible effects on earthworms of an extensive management including one annual mowing without any fertilizer input. Similarly, Decaëns et al. (1997) observed no changes in species richness during succession of a grazed chalk grassland over 44 years. In the present study, the most common earthworm species in both meadow types were A. rosea and O. lacteum, followed by A. caliginosa in abandoned sites and L. rubellus in managed sites. This is in line with the species assemblages of mountain grassland observed by Pop (1997). In the current study, managed meadows showed a marginally higher total earthworm density compared to abandoned ones, which is in contrast to other findings (Decaëns et al., 1997; Józefowska et al., 2018; Pižl, 1992). These contrasting findings let us assume that several context-specific factors such as species characteristics of the investigated plant and earthworm communities or ecosystem characteristics alter these ecological interactions. Earthworm abundance may be reduced via a decrease in nutrient supply and primary production due to changes in the floral composition (Scheu, 1992; Zaller and Arnone III, 1999a). Density of endogeic earthworms was marginally higher in managed meadows. Similarly, Ponge et al. (2013) found endogeic earthworms as K-selected species to be better adapted to disturbances than r-selected anecic species with a higher sensitivity. This may emphasize the marginally higher density of endogeic earthworms in managed sites, which are disturbed by mowing (Ivask et al., 2012). As soil properties between the abandoned and managed meadows were similar, it can be suggested that characteristics of vegetation may have caused the differences in earthworm populations. The marginally higher earthworm density in managed meadows may be the result of a higher plant species richness. Other studies reveal increasing earthworm density and biomass with higher plant species richness (Spehn et al., 2000; Zaller and Arnone III, 1999b). A loss of plant species leads to changes in community fine root biomass and negatively affects the food supply for earthworms (Zaller and Arnone III, 1999b). However, different earthworm densities in managed and abandoned meadows might have also affected root growth and aboveground plant production (Arnone III and Zaller, 2014). Furthermore, Eisenhauer et al. (2009) found the loss of key plant functional groups to be more important than plant species richness in shaping the structure of earthworm communities. Indeed, specific plant traits of functional groups appear to have a considerable influence on earthworm community (Piotrowska et al., 2013). Grasses may reduce the abundance of earthworms primarily due to their dense root system in plant communities (Eisenhauer et al., 2009; Piotrowska et al., 2013).
Considering interactions between earthworms and litter decomposition we observed contrasting patterns, i.e. less litter decomposition in mown meadows where more earthworms were present. Earthworms influence soil microbial communities and this affects microbial processes of soil organic matter and nutrient dynamics (Lavelle et al., 1998), and earthworms are considered important drivers of decomposition (Milcu et al., 2008; Seeber et al., 2006). Also, Lubbers et al. (2017) found earthworms to stimulate the decomposition of freshly added and older organic material rather than stabilize carbon inside biogenic aggregates. We explain our current contrasting findings with the method used to measure litter decomposition. Earthworms cannot pass through the mesh of the tea bags; thus only the potential influence of earthworms on microorganisms and mesofauna can be measured by tea bags (Brown, 1995; Wurst et al., 2018). Therefore, in order to identify which soil organisms affected litter decomposition in our meadows, more classical approaches to assess litter decomposition with different mesh sizes would be necessary (Hättenschwiler et al., 2005).
Our findings showed that an extensive management consisting of one annual mowing without fertilization of mountain meadows significantly increased the diversity of plant communities but did not affect the diversity of earthworm communities compared to leaving these meadows abandoned. Extensive meadow management also led to a reduced aboveground plant biomass production, increased earthworm densities and reduced litter decomposition. Aside from these contrasting effects on these parameters of supporting ecosystem services, effects on cultural ecosystem services such as health benefits of more diverse, extensively managed meadows have been shown (Arnberger et al., 2018). As this is one of few studies considering effects of extensive mountain meadow management on above- and belowground ecosystem interactions, more studies investigating these aspects seem imperative. Based on our study it can be recommended that a mix of abandoned and extensively managed meadows seems important in order to sustain plant and earthworm biodiversity in the study region.
Data are available upon request (contact: Ines Jernej, ines.jernej@gmx.net).
The supplement related to this article is available online at: https://doi.org/10.5194/we-19-53-2019-supplement.
IJ conducted the field work and further analyses. AB supported IJ in identifying plant species. IJ wrote the manuscript; RW and RIH assisted IJ with the field work and gave statistical advice. Project leaders JGZ, AA and TF designed and developed the “Healthy Alps” project. All authors reviewed the manuscript.
The authors declare that they have no conflict of interest.
We express our thanks to Johannes Karrer for providing support and the evaluation scheme. Special thanks are owed to the farmers and land owners for their permission to carry out the studies on their land.
This research has been supported by the Austrian Academy of Sciences (project: 10470 Healthy Alps). IJ was partly funded by Verband der Naturparke Österreichs and Österreichischen Bundesforste (research fund 2018/2019).
This paper was edited by Jutta Stadler and reviewed by two anonymous referees.
Arnberger, A., Eder, R., Allex, B., Ebenberger, M., Hutter, H.-P., Wallner, P., Bauer, N., Zaller, J. G., and Frank, T.: Health-related effects of short stays at mountain meadows , a river and an urban site – results from a field experiment, Int. J. Environ. Res. Public Health, 15, 1–19, https://doi.org/10.3390/ijerph15122647, 2018.
Arnone III, J. A. and Zaller, J. G.: Earthworm effects on native grassland root system dynamics under natural and increased rainfall, Front. Plant Sci., 5, 1–9, https://doi.org/10.3389/fpls.2014.00152, 2014.
Baur, B., Cremene, C., Groza, G., Rakosy, L., Schileyko, A. A., Baur, A., Stoll, P., and Erhardt, A.: Effects of abandonment of subalpine hay meadows on plant and invertebrate diversity in Transylvania, Romania, Biol. Conserv., 132, 261–273, https://doi.org/10.1016/j.biocon.2006.04.018, 2006.
Blouin, M., Hodson, M. E., Delgado, E. A., Baker, G., Brussaard, L., Butt, K. R., Dai, J., Dendooven, L., Peres, G., Tondoh, J. E., Cluzeau, D., and Brun, J. J.: A review of earthworm impact on soil function and ecosystem services, Eur. J. Soil Sci., 64, 161–182, https://doi.org/10.1111/ejss.12025, 2013.
Bobbink, R. and Willems, J. H.: Increasing dominance of Brachypodium pinnatum (L.) Beauv. in chalk grassland. A Threat to a species-rich Ecosystem, Biol. Conserv., 40, 301–314, https://doi.org/10.1016/0006-3207(87)90122-4, 1987.
Bobbink, R., During, H. J., Schreurs, J., Willems, J., and Zielman, R.: Effects of Selective Clipping and Mowing Time on Species Diversity in Chalk Grassland, Folia Geobot. Phytotx., 22, 363–376, 1987.
Bohner, A., Grims, F., Sobotik, M., and Zechner, L.: Die Trespen-Halbtrockenrasen (Mesobrometum erecti KOch 1926) im mittleren steirischen Ennstal (Steiermark, Österreich) – Ökologie, Soziologie und Naturschutz, Tuexenia, 23, 199–226, 2003.
Bohner, A., Starlinger, F., and Koutecky, P.: Vegetation changes in an abandoned montane grassland, compared to changes in a habitat with low-intensity sheep grazing – A case study in Styria, Austria, Eco. Mont., 4, 5–12, https://doi.org/10.1553/eco.mont-4-2s5, 2012.
Bouché, M. B.: Stratégies lombriciennes, in: Soil Oranism as Components of Ecosystems, Ecol. Bull., 25, 122–132, 1977.
Braun-Blanquet, J.: Pflanzensoziologie, 3rd edn., Springer-Verlag, Wien, 1964.
Brohmer, P.: Fauna von Deutschland, Ein Bestimmungsbuch unserer heimischen Tierwelt, Quelle & Meyer, Heidelberg, 1984.
Brown, G. G.: How do earthworms affect microflora and faunal community diversity?, Plant Soil, 170, 209–231, 1995.
Christian, E. and Zicsi, A.: Ein synoptischer bestimmungsschlussel der regenwurmer Osterreichs (Oligochaeta: Lumbricidae), Die Bodenkultur, 50, 121–131, https://doi.org/10.1002/anie.200805838, 1999.
Cluzeau, D., Guernion, M., Chaussod, R., Martin-Laurent, F., Villenave, C., Cortet, J., Ruiz-Camacho, N., Pernin, C., Mateille, T., Philippot, L., Bellido, A., Rougé, L., Arrouays, D., Bispo, A., and Pérès, G.: Integration of biodiversity in soil quality monitoring: Baselines for microbial and soil fauna parameters for different land-use types, Eur. J. Soil Biol., 49, 63–72, https://doi.org/10.1016/j.ejsobi.2011.11.003, 2012.
Decaëns, T., Dutoit, T., and Alard, D.: Earthworm community characteristics during afforestation of abandoned chalk grasslands (Upper Normandy, France), Eur. J. Soil Biol., 33, 1–11, 1997.
Del-Val, E. and Crawley, M. J.: What limits herb biomass in grasslands: Competition or herbivory?, Oecologia, 142, 202–211, https://doi.org/10.1007/s00442-004-1719-8, 2005.
Eisenhauer, N., Milcu, A., Sabais, A. C. W., Bessler, H., Weigelt, A., Engels, C., and Scheu, S.: Plant community impacts on the structure of earthworm communities depend on season and change with time, Soil Biol. Biochem., 41, 2430–2443, https://doi.org/10.1016/j.soilbio.2009.09.001, 2009.
Facelli, J. M. and Pickett, T. A.: Plant litter: Itsdynamics and effects on plant communitystructure, Bot. Rev., 57, 1–32, 1991.
Fischer, M. A., Oswald, K., and Adler, W.: Exkursionsflora für Österreich, Lichtenstein und Südtirol, 3rd edn., Land Oberösterreich, Biologiezentrum der Oberösterreichischen Landesmuseen, Linz, 2008.
Fontana, V., Radtke, A., Walde, J., Tasser, E., Wilhalm, T., Zerbe, S., and Tappeiner, U.: What plant traits tell us: Consequences of land-use change of a traditional agro-forest system on biodiversity and ecosystem service provision, Agric. Ecosyst. Environ., 186, 44–53, https://doi.org/10.1016/j.agee.2014.01.006, 2014.
Grime, J. P.: Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory, Am. Nat., 111, 1169–1194, https://doi.org/10.1086/283244, 1977.
Groffman, P. M., Eagan, P., Sullivan, W. M., and Lemunyon, J. L.: Grass species and soil type effects on microbial biomass and activity, Plant Soil, 183, 61–67, https://doi.org/10.1007/BF02185565, 1996.
Hättenschwiler, S., Tiunov, A. V., and Scheu, S.: Biodiversity and litter decomposition in terrestrial ecosystems, Annu. Rev. Ecol. Evol. Syst., 36, 191–218, https://doi.org/10.1146/annurev.ecolsys.36.112904.151932, 2005.
Hejcman, M., Hejcmanová, P., Pavlů, V., and Beneš, J.: Origin and history of grasslands in Central Europe – A review, Grass Forage Sci., 68, 345–363, https://doi.org/10.1111/gfs.12066, 2013.
Hooper, D. U., III, F. S. C., Ewel, J. J., Hector, A., Inchausti, P., Lavorel, S., Lawton, J. H., Lodge, D. M., Loreau, M., Naeem, S., Schmid, B., Setälä, H., Symstad, A. J., Vandermeer, J., and Wardle, D. A.: Effects of biodiversity on ecosystem functioning: a consensus of current knowledge, Ecol. Monogr., 75, 3–35, https://doi.org/10.1890/04-0922, 2005.
Hussain, R. I., Walcher, R., Brandl, D., Jernej, I., Arnberger, A., Zaller, J. G., and Frank, T.: Influence of abandonment on syrphid assemblages in mountainous meadows, J. Appl. Entomol., 142, 450–456, https://doi.org/10.1111/jen.12482, 2018.
Ivask, M., Kuu, A., Truu, M., Kutti, S., Meriste, M., and Peda, J.: Earthworm communities in soils of Estonian wooded meadows, Balt. For., 18, 111–118, 2012.
Jacquemyn, H., Brys, R., and Hermy, M.: Short-term effects of different management regimes on the response of calcareous grassland vegetation to increased nitrogen, Biol. Conserv., 111, 137–147, https://doi.org/10.1016/S0006-3207(02)00256-2, 2003.
Józefowska, A., Zaleski, T., Zarzycki, J., and Frączek, K.: Do mowing regimes affect plant and soil biological activity in the mountain meadows of Southern Poland?, J. Mt. Sci., 15, 2409–2421, https://doi.org/10.1007/s11629-018-4953-y, 2018.
Karrer, J.: Abandonment of nutrient poor montane meadows: impacts on regulating ecosystem services (decomposition, greenhouse gas efflux) and plant diversity, University of Natural Resources and Life Sciences, Vienna, 2015.
Kelemen, A., Török, P., Valkó, O., Miglécz, T. and Tóthmérész, B.: Mechanisms shaping plant biomass and species richness: Plant strategies and litter effect in alkali and loess grasslands, J. Veg. Sci., 24, 1195–1203, doi:10.1111/jvs.12027, 2013.
Kelemen, A., Török, P., Valkó, O., Deák, B., Miglécz, T., Tóth, K., Ölvedi, T., and Tóthmérész, B.: Sustaining recovered grasslands is not likely without proper management: vegetation changes after cessation of mowing, Biodivers. Conserv., 23, 741–751, https://doi.org/10.1007/s10531-014-0631-8, 2014.
Keuskamp, J. A., Dingemans, B. J. J., Lehtinen, T., Sarneel, J. M., and Hefting, M. M.: Tea Bag Index: a novel approach to collect uniform decomposition data across ecosystems, Methods Ecol. Evol., 4, 1070–1075, https://doi.org/10.1111/2041-210X.12097, 2013.
Klotz, S., Kühn, I., and Durka, W.: BIOLFLOR-Eine Datenbank zu biologisch-ökologischen Merkmalen der Gefäßpflanzen in Deutschland, Schriftenr. für Veg., 2002.
Köhler, B., Gigon, A., Edwards, P. J., Krüsi, B., Langenauer, R., Lüscher, A., and Ryser, P.: Changes in the species composition and conservation value of limestone grasslands in Northern Switzerland after 22 years of contrasting managements, Perspect. Plant Ecol. Evol. Syst., 7, 51–67, https://doi.org/10.1016/j.ppees.2004.11.003, 2005.
Lavelle, P.: Biology and ecology of earthworms, 3rd edn., Chapman & Hall, UK, 1997.
Lavelle, P. and Spain, A. V.: Soil Ecology, Kluwer Academic Publisher, Netherlands, 2001.
Lavelle, P., Pashanasi, B., Charpentier, F., Gilot, C., Rossi, J.-P., Derouard, L., Andre, J., Ponge, J.-F., and Bernier, N.: Large-Scale Effects of Earthworms on Soil Organic Matter and Nutrient Dynamics, in: Earthworm Ecology, edited by: C. A. Edwards, St. Lucie Press, 103–122, 1998.
Lieskovský, J., Bezák, P., Špulerová, J., Lieskovský, T., Koleda, P., Dobrovodská, M., Bürgi, M., and Gimmi, U.: The abandonment of traditional agricultural landscape in Slovakia – Analysis of extent and driving forces, J. Rural Stud., 37, 75–84, https://doi.org/10.1016/j.jrurstud.2014.12.007, 2015.
Liu, L., King, J. S., Booker, F. L., Giardina, C. P., Lee Allen, H., and Hu, S.: Enhanced litter input rather than changes in litter chemistry drive soil carbon and nitrogen cycles under elevated CO2: a microcosm study, Glob. Chang. Biol., 15, 441–453, https://doi.org/10.1111/j.1365-2486.2008.01747.x, 2009.
Lubbers, I. M., Pulleman, M. M., and Van Groenigen, J. W.: Can earthworms simultaneously enhance decomposition and stabilization of plant residue carbon?, Soil Biol. Biochem., 105, 12–24, https://doi.org/10.1016/j.soilbio.2016.11.008, 2017.
Lundber, A., Kapfer, J., and Måren, I. E.: Reintroduced mowing can counteract biodiversity loss in abandoned meadows, Erdkunde, 71, 127–142, 2017.
MacDonald, D., Crabtree, J. R., Wiesinger, G., Dax, T., Stamou, N., Fleury, P., Gutierrez Lazpita, J., and Gibon, A.: Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response, J. Environ. Manage., 59, 47–69, https://doi.org/10.1006/jema.1999.0335, 2000.
Marini, L., Klimek, S., and Battisti, A.: Mitigating the impacts of the decline of traditional farming on mountain landscapes and biodiversity: a case study in the European Alps, Environ. Sci. Policy, 14, 258–267, https://doi.org/10.1016/j.envsci.2010.12.003, 2011.
Maurer, K.: Natural and anthropogenic determinants of biodiversity of grasslands in the Swiss Alps, Universität Basel, 2005.
Meier, C. L. and Bowman, W. D.: Links between plant litter chemistry, species diversity, and below-ground ecosystem function, P. Natl. Acad. Sci. USA, 105, 19780–19785, https://doi.org/10.1073/pnas.0805600105, 2008.
Milcu, A., Partsch, S., Scherber, C., Weisser, W. W., and Scheu, S.: Earthworms and legumes control litter decomposition in a plant diversity gradient, Ecology, 89, 1872–1882, https://doi.org/10.1890/07-1377.1, 2008.
Moog, D., Poschlod, P., Kahmen, S., and Schreiber, K. F.: Comparison of species composition between different grassland management treatments after 25 years, Appl. Veg. Sci., 5, 99–106, https://doi.org/10.1111/j.1654-109X.2002.tb00539.x, 2002.
Mucina, L., Grabherr, G., and Ellmauer, T.: Die Pflanzengesellschaften Österreichs. Teil I: Anthropogene Vegetation, Gustav Fischer Verlag, Jena, 1993.
Niedrist, G., Tasser, E., Lüth, C., Dalla Via, J., and Tappeiner, U.: Plant diversity declines with recent land use changes in European Alps, Plant Ecol., 202, 195–210, https://doi.org/10.1007/s11258-008-9487-x, 2009.
Novara, A., Rühl, J., La Mantia, T., Gristina, L., La Bella, S., and Tuttolomondo, T.: Litter contribution to soil organic carbon in the processes of agriculture abandon, Solid Earth, 6, 425–432, https://doi.org/10.5194/se-6-425-2015, 2015.
Pavlů, L., Pavlů, V., Gaisler, J., Hejcman, M., and Mikulka, J.: Effect of long-term cutting versus abandonment on the vegetation of a mountain hay meadow (Polygono-Trisetion) in Central Europe, Flora – Morphol. Distrib. Funct. Ecol. Plants, 206, 1020–1029, https://doi.org/10.1016/j.flora.2011.07.008, 2011.
Peco, B., Carmona, C. P., de Pablos, I., and Azcárate, F. M.: Effects of grazing abandonment on functional and taxonomic diversity of Mediterranean grasslands, Agric. Ecosyst. Environ., 152, 27–32, https://doi.org/10.1016/j.agee.2012.02.009, 2012.
Piotrowska, K., Connolly, J., Finn, J., Black, A., and Bolger, T.: Evenness and plant species identity affect earthworm diversity and community structure in grassland soils, Soil Biol. Biochem., 57, 713–719, https://doi.org/10.1016/j.soilbio.2012.06.016, 2013.
Pižl, V.: Succession of earthworm populations in abandoned fields, Soil Biol. Biochem., 24, 1623–1628, https://doi.org/10.1016/0038-0717(92)90160-Y, 1992.
Ponge, J.-F., Pérès, G., Guernion, M., Ruiz-Camacho, N., Cortet, J., Pernin, C., Villenave, C., Chaussod, R., Martin-Laurent, F., Bispo, A., and Cluzeau, D.: The impact of agricultural practices on soil biota: A regional study, Soil Biol. Biochem., 67, 271–284, https://doi.org/10.1016/j.soilbio.2013.08.026, 2013.
Pop, V. V: Earthworm-Vegetation-Soil Relationships in the Romanian Carpathians, Soil Biol. Biochem., 29(, 223–229, 1997.
Prévosto, B., Kuiters, L., Bernhardt-Römermann, M., Dölle, M., Schmidt, W., Hoffmann, M., Van Uytvanck, J., Bohner, A., Kreiner, D., Stadler, J., Klotz, S., and Brandl, R.: Impacts of Land Abandonment on Vegetation: Successional Pathways in European Habitats, Folia Geobot., 46, 303–325, https://doi.org/10.1007/s12224-010-9096-z, 2011.
R Core Team: A language and environment for statistical computing, Version v 3.1., Vienna, Austria, available at: http://www.R-project.org (last access: 23 August 2019), 2016.
Rolecek, J., Cornej, I. I., and Tokarjuk, A. I.: Understanding the extreme species richness of semi-dry grasslands in east-central Europe: A comparative approach, Preslia, 86, 13–34, 2014.
Scherer-Lorenzen, M.: Functional diversity affects decomposition processes in experimental grasslands, Funct. Ecol., 22, 547–555, https://doi.org/10.1111/j.1365-2435.2008.01389.x, 2008.
Scheu, S.: Changes in the lumbricid coenosis during secondary succession from a wheat field to a beechwood on limestone, Soil Biol. Biochem., 24, 1641–1646, https://doi.org/10.1016/0038-0717(92)90163-R, 1992.
Schmitzberger, I., Wrbka, T., Steurer, B., Aschenbrenner, G., Peterseil, J., and Zechmeister, H. G.: How farming styles influence biodiversity maintenance in Austrian agricultural landscapes, Agric. Ecosyst. Environ., 108, 274–290, https://doi.org/10.1016/j.agee.2005.02.009, 2005.
Seeber, J., Scheu, S., and Meyer, E.: Effects of macro-decomposers on litter decomposition and soil properties in alpine pastureland: A mesocosm experiment, Appl. Soil Ecol., 34, 168–175, https://doi.org/10.1016/j.apsoil.2006.02.004, 2006.
Spehn, E. M., Joshi, J., Schmid, B., Alphei, J., and Körner, C.: Plant diversity effects on soil heterotrophic activity in experimental grassland ecosystems, Plant Soil, 224, 217–230, https://doi.org/10.1023/A, 2000.
Tasser, E. and Tappeiner, U.: Impact of land use changes on mountain vegetation, Appl. Veg. Sci., 5, 173–184, https://doi.org/10.1658/1402-2001(2002)005[0173:IOLUCO]2.0.CO;2, 2002.
Walcher, R., Karrer, J., Sachslehner, L., Bohner, A., Pachinger, B., Brandl, D., Zaller, J. G., Arnberger, A., and Frank, T.: Diversity of bumblebees, heteropteran bugs and grasshoppers maintained by both: abandonment and extensive management of mountain meadows in three regions across the Austrian and Swiss Alps, Landsc. Ecol., 32, 1937–1951, https://doi.org/10.1007/s10980-017-0556-1, 2017.
Wehn, S., Taugourdeau, S., Johansen, L., and Hovstad, K. A.: Effects of abandonment on plant diversity in semi-natural grasslands along soil and climate gradients, J. Veg. Sci., 28, 838–847, https://doi.org/10.1111/jvs.12543, 2017.
Weiss, L. and Jeltsch, F.: The response of simulated grassland communities to the cessation of grazing, Ecol. Modell., 303, 1–11, https://doi.org/10.1016/j.ecolmodel.2015.02.002, 2015.
Wilson, J. B., Peet, R. K., Dengler, J., and Pärtel, M.: Plant species richness: The world records, J. Veg. Sci., 23, 796–802, https://doi.org/10.1111/j.1654-1103.2012.01400.x, 2012.
Wurst, S., Sonnemann, I., and Zaller, J. G.: Soil Macro-Invertebrates: Their Impact on Plants and Associated Aboveground Communities in Temperate Regions BT – Aboveground–Belowground Community Ecology, in Aboveground-Belowground Community Ecology, edited by: Ohgushi, T., Wurst, S., and Johnson, S. N., Springer International Publishing, 175–200, 2018.
Xiong, S. and Nilsson, C.: The effects of plant litter on vegetation: a meta-analys, J. Ecol., 87, 984–994, 1999.
Zaller, J. G. and Arnone III, J. A.: Earthworm and soil moisture effects on the productivity and structure of grassland communities, Soil Biol. Biochem., 31, 517–523, 1999a.
Zaller, J. G. and Arnone III, J. A.: Earthworm responses to plant species' loss and elevated CO2 in calcareous grassland, Plant Soil, 208, 1–8, https://doi.org/10.1023/A:1004424720523, 1999b.
Zentralanstalt für Meteorologie und Geodynamik (ZAMG): Klimamittel – ZAMG, 2018.