the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 15 Sep 2025
| 15 Sep 2025
Introducing a glacier forefield monitoring site network to understand succession in the Northern Limestone Alps
Christian Hecht
Ulrike Herzschuh
Dirk Scherler
Since the end of the Little Ice Age (ca. 1855), glaciers retreated in the Alps, leaving new ground for genuine primary succession. The patterns and processes of glacier forefield succession have been studied globally for decades. Surprisingly, no such analysis exists from the Northern Limestone Alps. We therefore initiated a monitoring scheme with permanent plots to study plant succession and vegetation assembly at four forefields, namely the Hallstätter Glacier, Großer Gosau Glacier (both at Dachstein massif, Austria), Watzmann Glacier, and Blaueis (both at Berchtesgaden National Park, Germany), which is abbreviated as the BDGF (Berchtesgaden-Dachstein Glacier Forefield) platform. The aim of the long-term research envisaged and performed in this platform is to get a better understanding of the vegetation succession and community assembly in the glacier forefield development of the Northern Limestone Alps, using a multidisciplinary approach. Here, we introduce the basic characteristics of the BDGF platform; i.e. we describe the monitoring network, the observational design, and the methodological approaches. We present the baseline vegetation characteristics, and we outline the studies already initiated or to be performed in the near future. The methodology encompasses a chronosequence approach, where plots, using a frequency grid frame of 1 m × 1 m, are placed in specific successional stages (related to age classes since deglaciation). We show that, as expected, species richness and cover increase with age. Unexpectedly, though, these processes seem to be much slower than what has been observed in the Central Alps on siliceous substrates. We suggest that this could be due to the geological substrate, i.e. its chemistry as well as its karstic conditions, but also due to the morphology of the terrain, which hardly enables species colonization from above (i.e. following gravity) but mainly from below.
- Article
(9277 KB) - Full-text XML
- BibTeX
- EndNote




The Alpine glaciers reached their largest extent in the mid-1850s. Since then, they retracted gradually, although there were some less pronounced advances in the 20th century (Kuhn et al., 1997; Mayer, 2012). These, however, differed locally. In the Northern Limestone Alps, the Watzmann glacier and Blaueis in the Berchtesgaden National Park had some larger extent in the 1920s (http://www.bayerische-gletscher.de/blaueis.htm , last access: 3 September 2025, Hagg, 2006). Some Dachstein glaciers (such as the Hallstätter Glacier) had some smaller progressions in the late 1920s and 1980s (Helfricht, 2009).
Retreating glaciers leave behind bare bedrock or unconsolidated sediments that were previously covered by ice for centuries to millennia (Nicolussi et al., 2022), which provide new opportunities for the succession of vegetation. These ecosystems, emerging from formerly glaciated areas, are delicate systems that deserve conservation efforts (Bosson et al., 2023), and which provide important grounds to study the ecosystem development and associated ecosystem functions (Losapio et al., 2025). Ecologists were hence interested in the patterns and processes of glacier forefield vegetation succession in the Alps as early as the 1880s in the Rhône Glacier in Valais, Switzerland (Coaz, 1887). The more systematic studies started in the 1950s with studies of the Schwarzensteinkees in the Zillertaler Alps (Frenzel, 1951/1957; Krause and Frenzel, 1997) and in the 1960s with the Gaisberg and Rotmoos Glacier in the Ötztal Alps (Jochimsen, 1963) (both Austria). These early studies, though, were hardly able to reconstruct the glacier forefield succession along dated sites, i.e. knowing at which point in time the glacier retreated. Since the pioneering work of Burga (1999) at the Morteratsch Glacier (Grison, Switzerland) in the 1980s and of Erschbamer et al. (1999, 2023) and Raffl et al. (2006a) at the Rotmoosferner (Ötztal, Austria) in the 1990s, a chronosequence approach was more systematically applied. This means that the study sites were placed in a systematic way, following the retreat of the glaciers, and are dated according to the year they were exposed.
Since then, numerous studies were published, studying plant succession of single glaciers in the central Italian Alps (e.g. Cannone et al., 2008; Cannone and Piccinelli, 2021) and the Southern and Central Alps across France, Italy, Switzerland, and Austria (see overview in the reviews of Cauvy-Fraunié and Dangles, 2019, and Ficetola et al., 2021). A common pattern is, obviously, the increase in species richness and cover with site age. The development of glacier forefields is schematically summarized by Eichel (2019), incorporating geomorphological processes and biotic processes into a biogeomorphic framework. Overall, it seems that in some regions the vegetation can recover as quickly as within 100 years to form dense Alpine swards and after 150 years being very close to the surrounding Alpine meadows or even showing shrub and tree succession (see Burga, 1999; Nagl and Erschbamer, 2010; Eichel, 2019; Eichel et al., 2023). Further, Hanusch et al. (2022) studied five organismal groups including plants, animals, and microbes, summarized as “multidiversity”. They showed that stochastic processes (related to dispersal and colonization) seem to be prevalent for the first ca. 60 years, whereas deterministic processes (such as biotic interactions) dominate in late successional stages. The change in plant community composition (total β diversity) was nicely quantified and partitioned in the elements of β richness and β replacement (following Carvalho et al., 2012) across the forelands of 46 retreating glaciers worldwide (18 in the European Alps) by Cantera et al. (2024).
It is striking, though, that studies on glacier forefield vegetation succession seemingly left out the Northern Alps, in particular the Northern Limestone Alps. There were some intense studies performed on the vegetation of the Dachstein range (e.g. by Grims, 1982; Moser and Vareschi, 1959). They classified different stages of vegetation succession, but unfortunately, their relevés can neither be geolocated properly nor dated with respect to age since deglaciation. Furthermore, we are not aware of other studies on other glacier forefields in the Northern Limestone Alps. This is even more striking, considering that the environmental conditions are highly specific. Weathering and chemical composition of the Dachstein limestone are different to those of the typical gneiss or granite substrates in the Central Alps. Specifically, the carbonate bedrock limits the formation of clay minerals, and in the karstic environments, nutrients, fine particles, and initial soil get easily washed away into the underground; water is scarce during summer. Furthermore, there are indications that vegetation on calcareous substrates responds differently to climate change than that on siliceous substrates (Nicklas et al., 2021). In addition, the Northern Limestone Alps glaciers have exclusively northern expositions and are much lower than those in the Central Alps.
To close this research gap, in 2016 we initiated a monitoring of glacier forefield vegetation, following a strict chronosequence approach. In this paper, our main aim is to introduce and describe the basic design of this monitoring network in the Northern Limestone Alps, specifically covering the Dachstein range (with Hallstätter Glacier and Großer Gosau Glacier) in Austria and Berchtesgaden National Park (with Watzmann Glacier and Blaueis) in Germany (the Berchtesgaden-Dachstein Glacier Forefield (BDGF) platform). Our broad interest is to study various aspects of glacier forefield succession. These encompass questions such as the following: how does the functional composition of vegetation change with glacier forefield age? What is the relation between geological (land surface processes) and biogenic processes during the succession? Can successional development from ground vegetation be related to palaeostratic patterns from lake sediment analyses? However, these are general questions for this platform. We additionally report the first results, specifically differences in species composition among glaciers, as well as changes in species richness, frequency, and cover with successional age. Lastly, we outline a future interdisciplinary research agenda planned on this platform. To easily refer to this monitoring platform, we use the acronym BDGF, i.e. Berchtesgaden-Dachstein glacier forefield. This paper shall serve as a baseline and reference for the follow-up studies.
2.1 Study areas
2.1.1 Locations
The BDGF is situated in two mountain ranges of the Northern Limestone Alps, namely the Dachstein Mountains and the Berchtesgaden Alps (Berchtesgaden National Park), with two glacier forefields as study sites, each (Fig. 1).
The Dachstein Mountains are located in northeast Austria (southern Upper Austria, eastern Salzburg, and northwestern Styria) with the Hoher Dachstein (2995 m a.s.l.) as the highest mountain. The two glacier forefields are beneath the Hallstätter Glacier (elevation of the plots range between ca. 2060 and 2300 m a.s.l.) and the Großer (Great) Gosau Glacier (ca. 2160–2520 m a.s.l.), north of the Hoher Dachstein.
The Berchtesgaden Alps are located in the most southeastern tip of Germany (Bavaria) and border Austria (Salzburg), while the Berchtesgaden National Park covers parts of the southernmost section of the Berchtesgaden Alps in Germany. The two glacier forefields are in the Watzmannkar (Watzmann cirque) beneath the (former) Watzmann Glacier (plots ca. 1970–2030 m a.s.l.), with an average elevation of 2060 m a.s.l., being the lowermost glacier in Bavaria (Mayer et al., 2021), and in the Blaueiskar of the Blaueis (ca. 1915–2035 m a.s.l.), the northernmost glacier of the Alps.
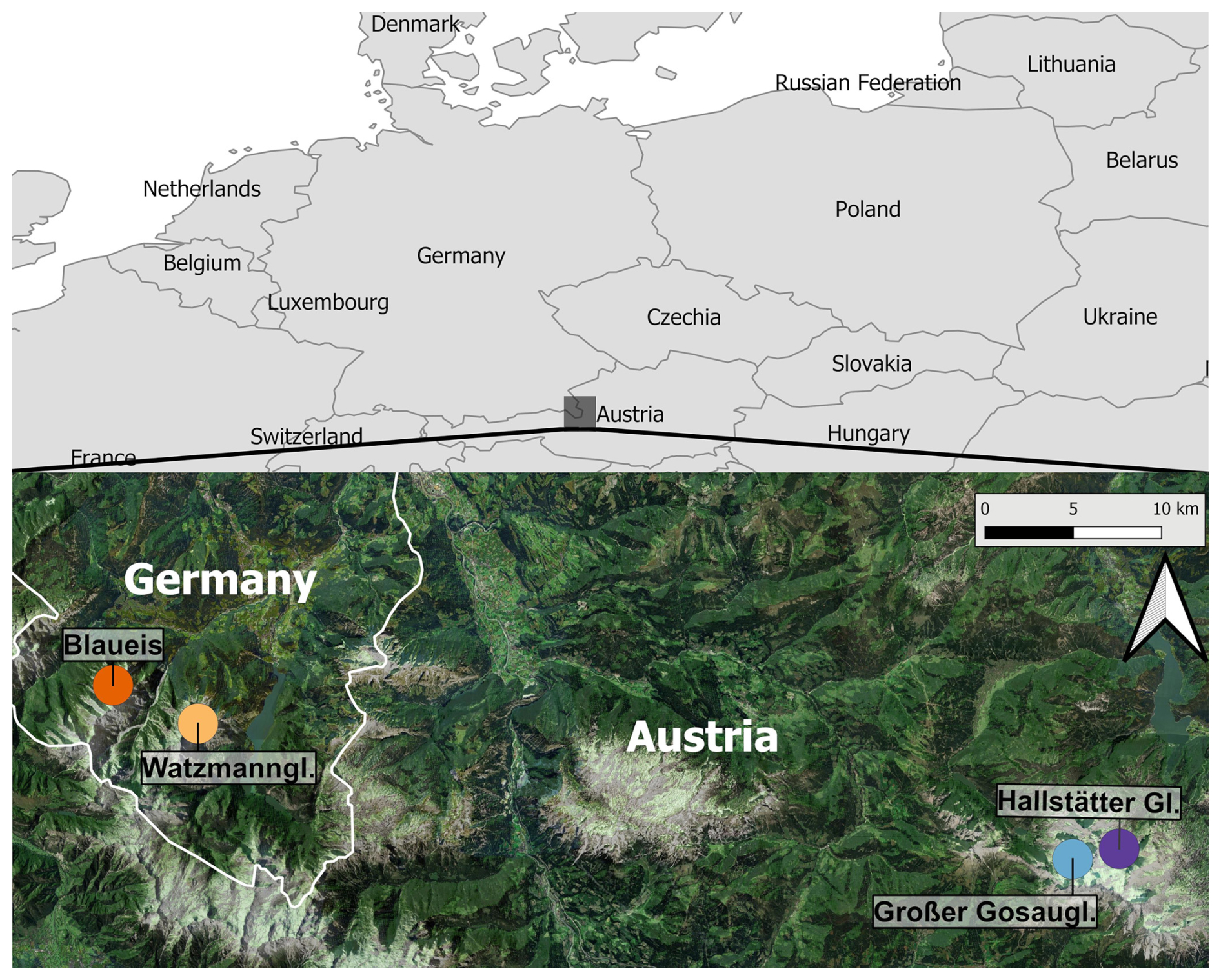
Figure 1Location of the study sites. Blaueis and Watzmann Glacier (Watzmanngletscher) are located in the Berchtesgaden Alps in Germany. Great Gosau Glacier (Großer Gosaugletscher) and Hallstätter Glacier (Hallstätter Gletscher) belong to the Dachstein Mountains in Austria. Map based on Geoland.at (2019).
2.1.2 Geology and geomorphology
The Dachstein Massif and the Berchtesgaden Alps are two groups of impressive peaks of the Northern Limestone Alps. Their bedrock is dominated by the prominent “Dachsteinkalk”, a series of banked limestones and dolomites that were deposited under shallow water conditions in the Tethyan Sea, ca. 200–240 million years ago. The strata have mostly gentle dips and are heavily karstified. The Dachstein limestones are weathering rather slowly under the cold Alpine conditions and result in alkaline conditions in the thin soils and protosoils (Ganss and Grünfelder, 1979; Moser, 1997; Scheidleder et al., 2001). This Dachstein limestone is prevailing in all four study regions. In most places, Pleistocene glaciation eroded the uppermost bedrock and stripped away surficial sediments. Present-day sedimentary cover corresponds to lateral and terminal moraines as well as basal till.
Common characteristics of all four glaciers are their relatively low slopes and wide extents, which are due to the plateau-like morphology of the Dachsteinkalk massifs. As a result, the glacier forefields are similarly broad and gentle sloping, mostly to the north.
2.1.3 Climate
The climate of the region is oceanic with Alpine influence and characterized by western and northern winds with high precipitation in summer and long snow cover in winter. Mean annual temperatures range between −2 and 2 °C (depending on elevation), with average annual minima between −20 and −24 °C and maxima between 18 and 22 °C. Annual precipitation ranges between 2500 and 3000 mm in the Dachstein region (CLARISIA, 2015). In the Berchtesgaden Alps, mean temperatures are more towards the upper limits of the aforementioned ranges, and annual precipitation tends towards the lower limits of the aforementioned ranges (https://klimainformationssystem.bayern.de, last access: 3 September 2025).
2.2 Plot layout and design
In each glacier forefield, we established 13 permanent plots, recorded GPS coordinates, and permanently marked them with a magnetic ring fixed to a 17 cm iron nail, driven into the ground at each corner of the plot. At the areas covering the different periods of deglaciation, i.e. around 1980, 1950, and at the terminus during the largest extent (ca. 1855 in the Dachstein region and ca. 1920 at the Berchtesgaden Mountains), we established 3 plots each, except close to the current glacier terminus at the time of establishment (less than ca. 10 years since deglaciation), where we established 4 plots to consider the more stochastic colonization during early succession (Hanusch et al., 2022). The extent of the glaciers at different times was available from https://www.doris.at/themen/umwelt/pdf/clairisa/karten/Gletscher_Stand.pdf (last access: 3 September 2025) and kindly provided as GIS shapefiles by Blue Sky Wetteranalysen, Attnang, Austria (https://www.blueskywetter.com, last access: 3 September 2025) for the Dachstein glaciers and from http://www.bayerische-gletscher.de (last access: 3 September 2025) for the Berchtesgaden glaciers.
When selecting plot locations, we considered the variability of conditions within each successional age class by placing the different plots as far as possible from each other (see Fig. 2). Further, we also avoided possible disturbances from humans or landslides so that ideally age since deglaciation equals successional age (see Wojcik et al., 2021), translating to successional stage (i.e. the state of vegetation development). Usually, plots farther away from the glacier terminus are also older (Hallstätter Glacier and Großer Gosau Glacier). Most glaciers, e.g. Hallstätter Glacier Großer Gosau Glacier, exhibit a considerable slope which usually correlates with the age of deglaciation (i.e. the older, the further away and downward from the glacier terminus). The forefield of the Watzmann Glacier, though, is located on terrain with very little downhill gradient but considerable microtopographical heterogeneity. Therefore, we assessed the ice extent relating the on-site topographic situation to ice cover data from http://www.bayerische-gletscher.de (last access: 3 September 2025). The Blaueis receives many visitors from the close-by Blaueishütte (mountain refuge of the German Alpine Club – DAV), so the ground below the glacier terminus is highly disturbed. Here, we placed most plots at different terrace levels above the (former) glacier.
In the Dachstein region, we established plots in 2016 and first sampled them from 2017, while in the Berchtesgaden National Park we established and first sampled them in 2018.
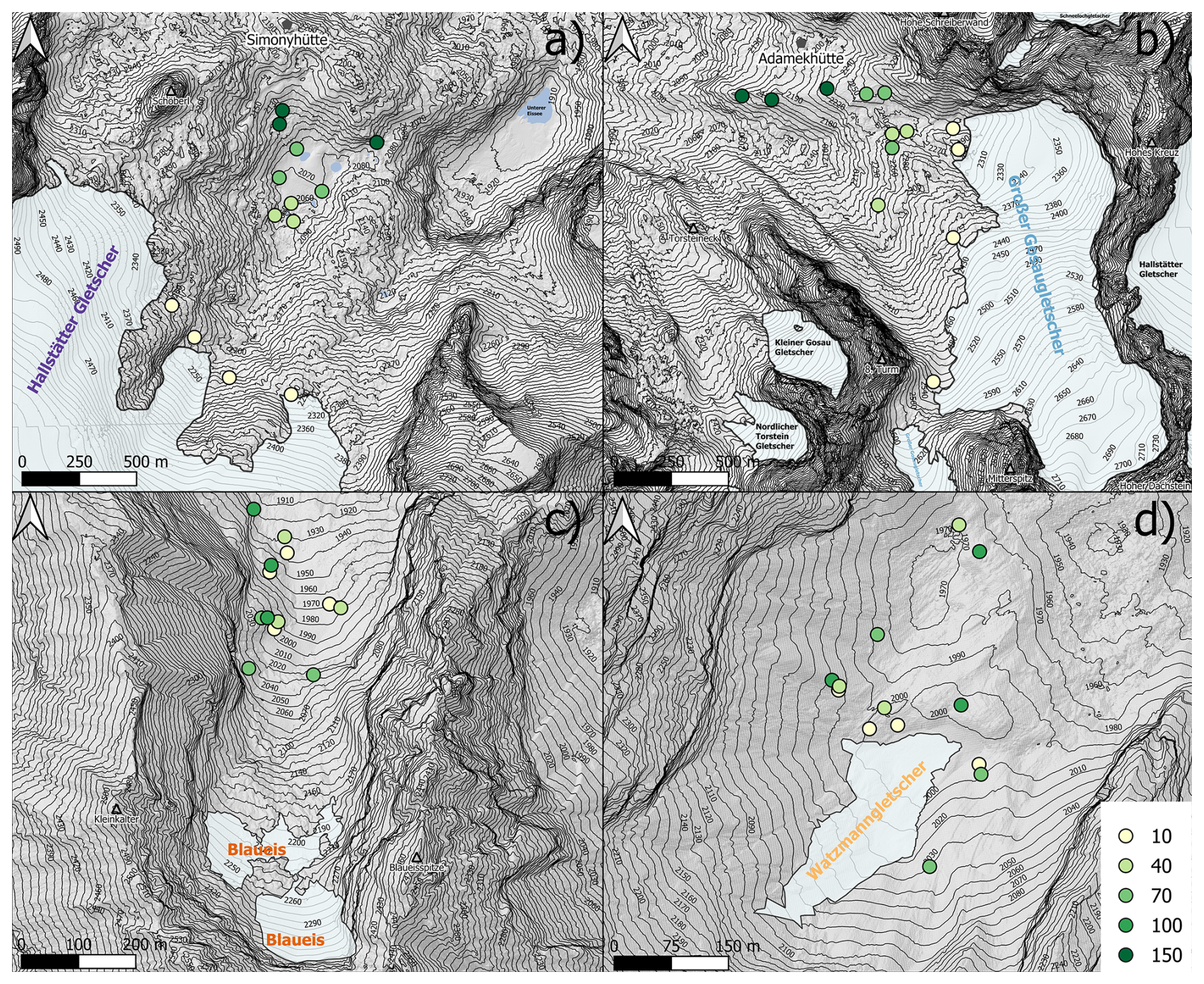
Figure 2Location of the plots in the forefields of Hallstätter Glacier (a), Großer Gosau Glacier (b), Blaueis (c), and Watzmann Glacier (d). The plots of the areas are colour-coded according to the age classes (10, 40, 70, 100, and 150 years since deglaciation). Digital elevation model for map creation was used from the regional government of Upper Austria (https://www.data.gv.at (last access: 3 September 2025) for Dachstein mountain and from the Bavarian Surveying Administration for Berchtesgaden https://geodaten.bayern.de/opengeodata (last access: 3 September 2025).
2.3 Vegetation survey methods
We survey the Dachstein glacier plots every odd year and those of the Berchtesgaden National Park every even year, both in late July to mid-August, which resulted in 52 plots sampled four times, each, until 2024. In the case of unfavourable weather or snow cover preventing sampling, we revisited the plots later in the season or the following year (with surveys in both regions, then).
At each plot, we placed a 1×1 m2 grid frame with one hundred 10×10 cm2 grid cells, separated by white strings, with the corners at the four magnetic nails and with the first row facing towards the glacier. Within each grid cell, we recorded all vascular species, as well as the presence of mosses and lichens, or whether that grid cell was completely covered by rocks and hence unavailable to be colonized by vascular plants. This resulted in a frequency per plant species per plot record. The method follows the 1 m2 subplot-frequency counts of the GLORIA protocol (Pauli et al., 2015). In addition, we estimated the total cover of all vascular plant species per plot in percent since 2022. Plant nomenclature follows Aeschimann et al. (2004).
2.4 Statistical methods
We used the baseline plant frequency data from 2017 (Dachstein glaciers) and 2018 (Berchtesgaden glacier) and cover data estimated in 2022 (all sites) for the analysis. We grouped the age of the plots (i.e. years since deglaciation) into the following classes: ca. 10, 40, 70, 100 (only Berchtesgaden), and 150 (only Dachstein) years. For all analyses, we used the statistical environment R (R Core Team, 2023), Version 4.3.0. To test for differences in species richness among the glaciers, we used a mixed effects analysis of variance (with glacier forefield nested in a mountain range as random effect). To test for changes in species richness, frequency, and total cover, we used mixed effects models with species richness or frequency, respectively, as response variable and the age class of the plots (since deglaciation) as predictor. Analogously, we modelled the frequency of species by using a generalized mixed effects model with binomial error. For these analyses, we started with a full model, using the interaction of age since deglaciation and glacier as predictors and then using a stepwise simplification according to error probability (i.e. removing the insignificant variables, starting with the interaction). Here, we only present the minimal adequate model results (Crawley, 2012). The nesting of plots in a glacier forefield in a mountain range was coded as a random effect, using the R functions lmer() and glmer() for species richness and for frequency and cover, respectively, in lme4 (V 1.1-35.3). We calculated error probabilities using lmerTest (V 3.1-3), ANOVA differences using car (V 3.1-2), and Tukey post hoc test with emmeans (V1.102). Finally, we explored patterns of variation between study areas, computing a principal coordinate analysis (PCoA, computed using wcmdscale()), using Bray–Curtis distance (function vegdist()) as a measure of dissimilarity (both using vegan (V 2.6-61)). Figures were created using ggplot (V 3.5.1).
3.1 Species composition
The species composition differed markedly between the forefields of Berchtesgaden and Dachstein Mountains, with higher similarities observed within the mountain ranges than between them (Fig. 3a). The first axis can be interpreted as representing the successional gradient. Here species such as Arabis alpina or Hornungia (Pritzelago) alpina are typical for earlier successional stages, while species such as Silene acaulis, dwarf willows (e.g. Salix serpyllifolia) or Carex firma represent later successional stages in the two Dachstein glacier forefields (both having considerable overlap in species composition). The second axis represents largely differences in species composition between the Dachstein and the Berchtesgaden glacier forefields (the latter with less of a compositional overlap between the two forefields compared to the former). Typical for Berchtesgaden glacier forefields are Thlaspi rotundifolia (almost exclusively found there) or Moehringia ciliata.
Calculating median species frequency per plot, the highest frequencies were in the forefields of the Hallstätter Glacier, followed by Blaueis and Great Gosau Glacier. The lowest frequencies were found in the forefield of the Watzmann Glacier (Fig. 3b–e). Accordingly, the most frequently recorded species across all areas differ among mountain ranges and forefields. Hornungia alpina is the one species among those most frequently found across all four glacier forefields. Others are Poa alpina and Cerastium uniflorum. The highest median frequencies (i.e. > 10) are reached by Campanula cochleariifolia, Hornungia alpina, Gentiana orbicularis, and Euphrasia minima in the Hallstätter glacier forefield. Those of the Great Gosau glacier forefield are only two species, namely Cerastium uniflorum and (again) Hornungia alpina. In the Blaueis forefield, only Hornungia alpina exceeds a median frequency of 10, while in the Watzmann glacier forefield, none of the three most common species is that frequent.
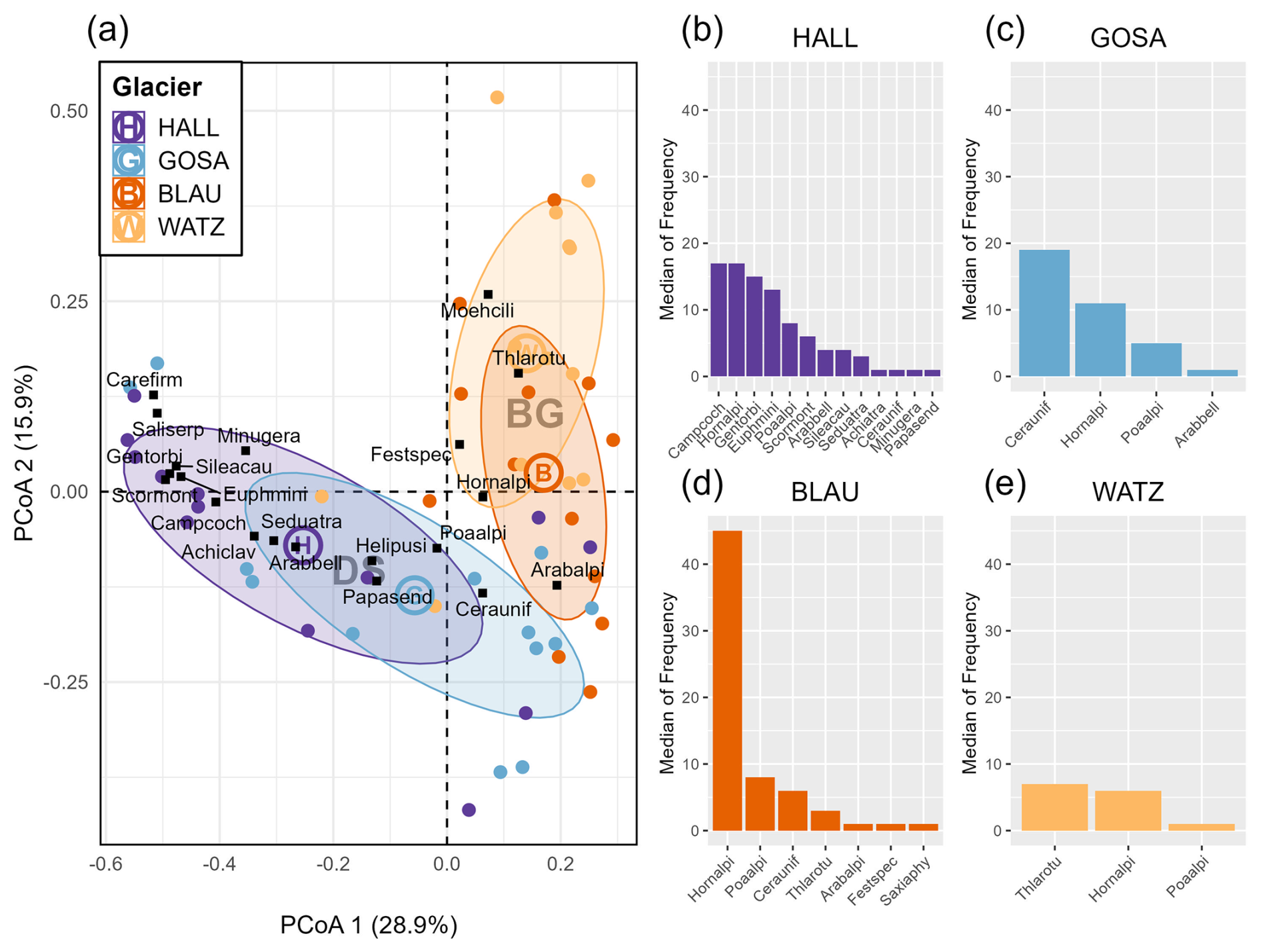
Figure 3(a) PCoA (principal coordinate analysis) for all plots of the study areas (HALL – Hallstätter Glacier, GOSA – Great Gosau Glacier, BLAU – Blaueis, WATZ – Watzmann Glacier), including the centroids of these areas and the corresponding mountain ranges (DS – Dachstein Mountain, BG – Berchtesgaden). The first 20 most frequent species across all study areas are shown in the PCoA. (b–e) Median of species frequency per plot for each glacier. Only species with median frequency > 0 are shown. Abbreviations of the plant species: Arabalpi – Arabis alpina, Arabbell – Arabis bellidifolia, Achiatra – Achillea atrata, Achiclav – Achillea clavennae, Campcoch – Campanula cochleariifolia, Carefirm– Carex firma, Ceraunif – Cerastium uniflorum, Euphmini – Euphrasia minima, Festspec – Festuca spec., Genorbi – Gentiana orbicularis, Helipusi – Heliosperma pusillum, Hornalpi – Hornungia alpina, Minugera – Minuartia gerardii, Moehcili – Moehringia ciliata, Papasend – Papaver alpinum ssp. sendtneri, Poaalpi – Poa alpina, Saliserp – Salix serpyllifolia, Saxiaphy – Saxifraga aphylla, Scormont – Scorzoneroides montana, Seduatra – Sedum atratum, Sileacau – Silene acaulis, Thlarotu – Thlaspi rotundifolium. Baseline data are from 2017 (Dachstein Mountains) and 2018 (Berchtesgaden Alps).
3.2 Species richness
Species numbers did not differ significantly among the glacier forefields (Table 1), although the highest median species richness was in the forefields of the Hallstätter Glacier (median = 14) and Great Gosau Glacier (9), followed by Blaueis (7) and lowest richness in the Watzmann cirque (3) (Fig. 4a). With an increase in age of the forefield plots (time since deglaciation), species numbers per plot across all study areas increased significantly (Fig. 4b, Table 2) across all glaciers. While the slopes did not differ significantly, their intercepts differed, corresponding to the order above.
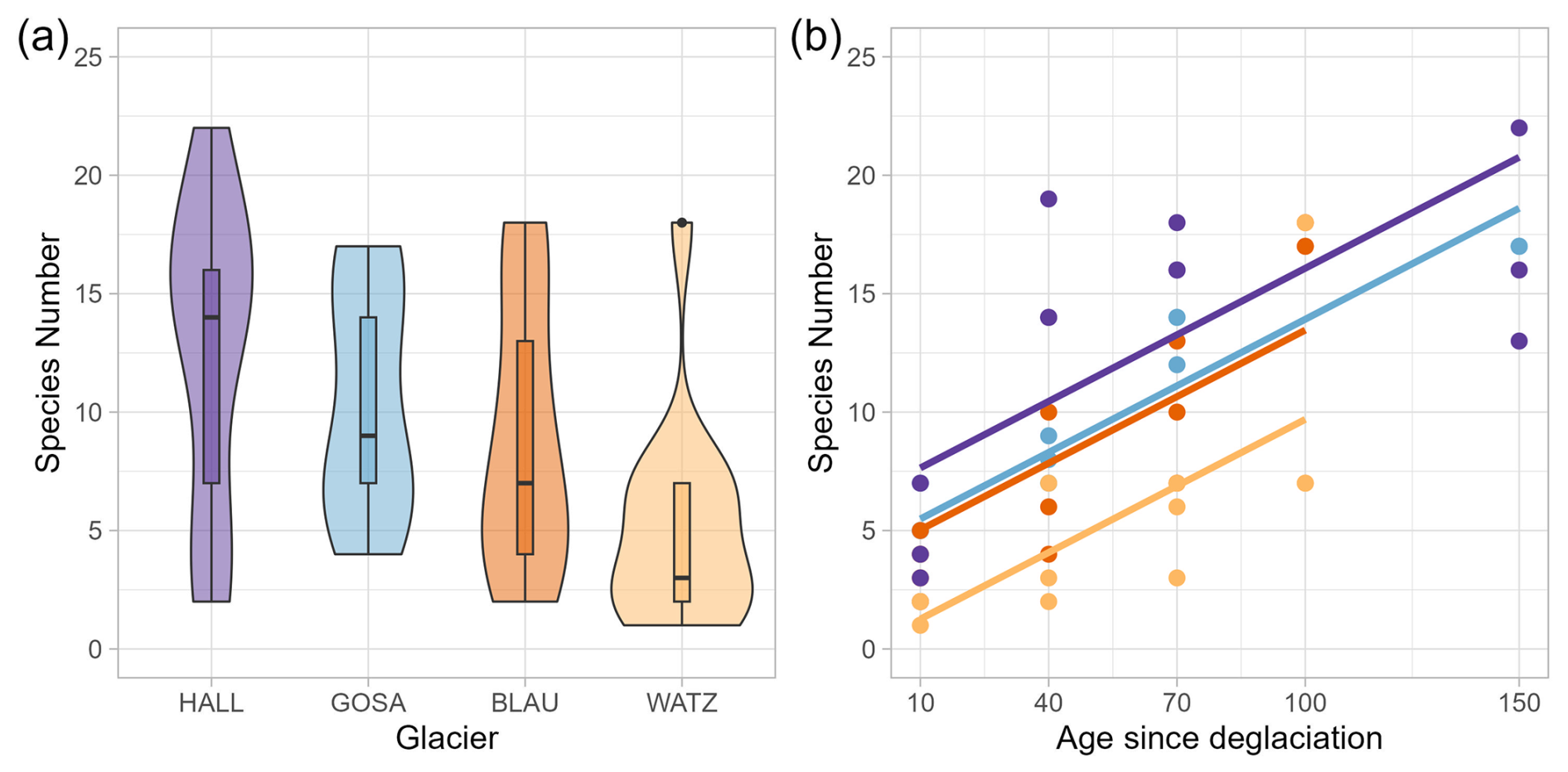
Figure 4(a) Species numbers for the four glaciers (HALL – Hallstätter Glacier, GOSA – Great Gosau Glacier, BLAU – Blaueis, WATZ – Watzmann Glacier; lines in boxes of violin plots represent medians; boxes represent the 25th to 75th percentiles); (b) species numbers of four successional stages (age since deglaciation) across all study areas. Baseline data from 2017 (Dachstein Mountains) and 2018 (Berchtesgaden Alps) were used.
Table 1Results of nested ANOVA of the species richness of the four glacier forefields with forefield nested in a mountain as random effect (corresponding to Fig. 4a).
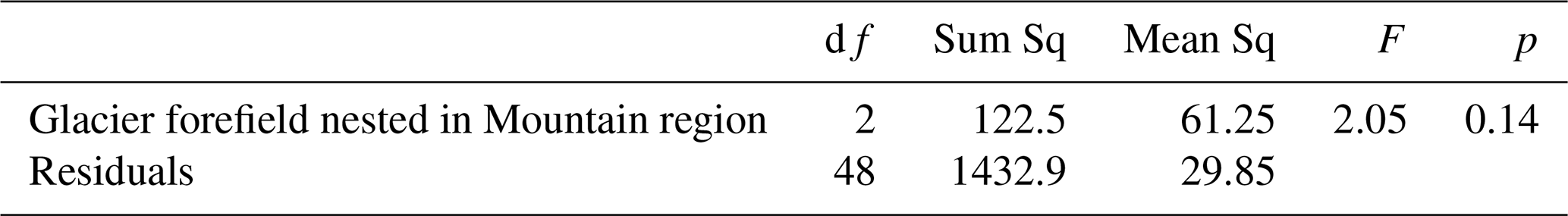
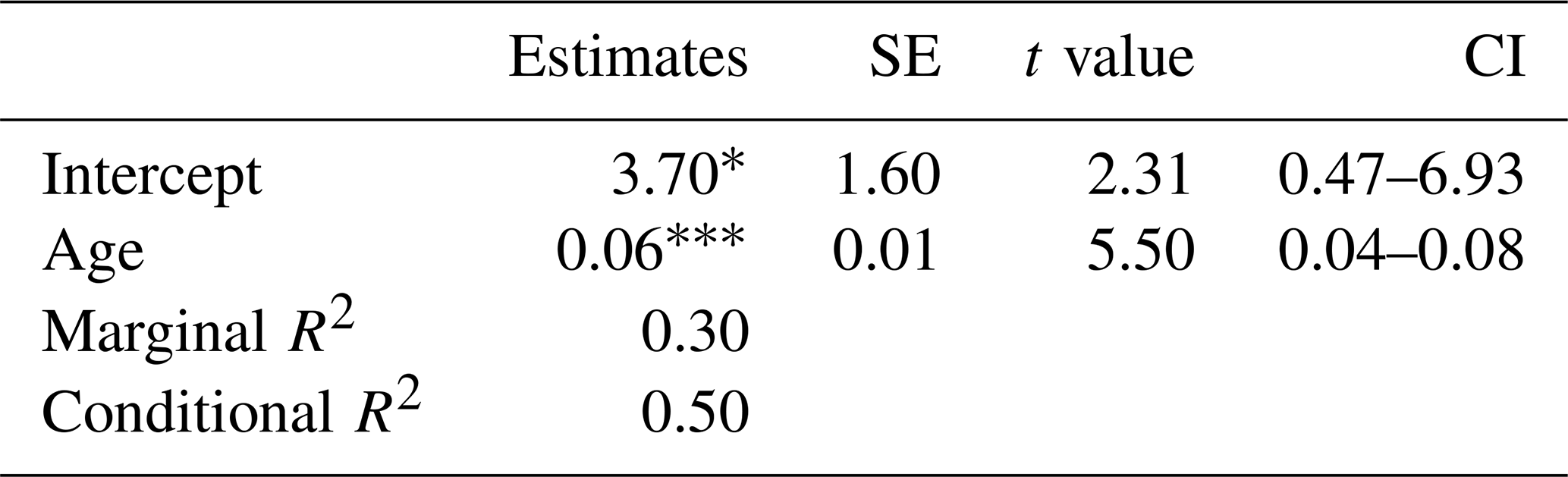
Table 2Summary statistics for the linear mixed effects model of species richness in response to age (time since deglaciation) and glacier forefield nested in a mountain as random effect (corresponding to Fig. 4b; see there for abbreviations), SE: standard error, CI: confidence interval. * 0.01 < p ≤ 0.05; p < 0.001.
3.3 Plant frequency and cover per plot
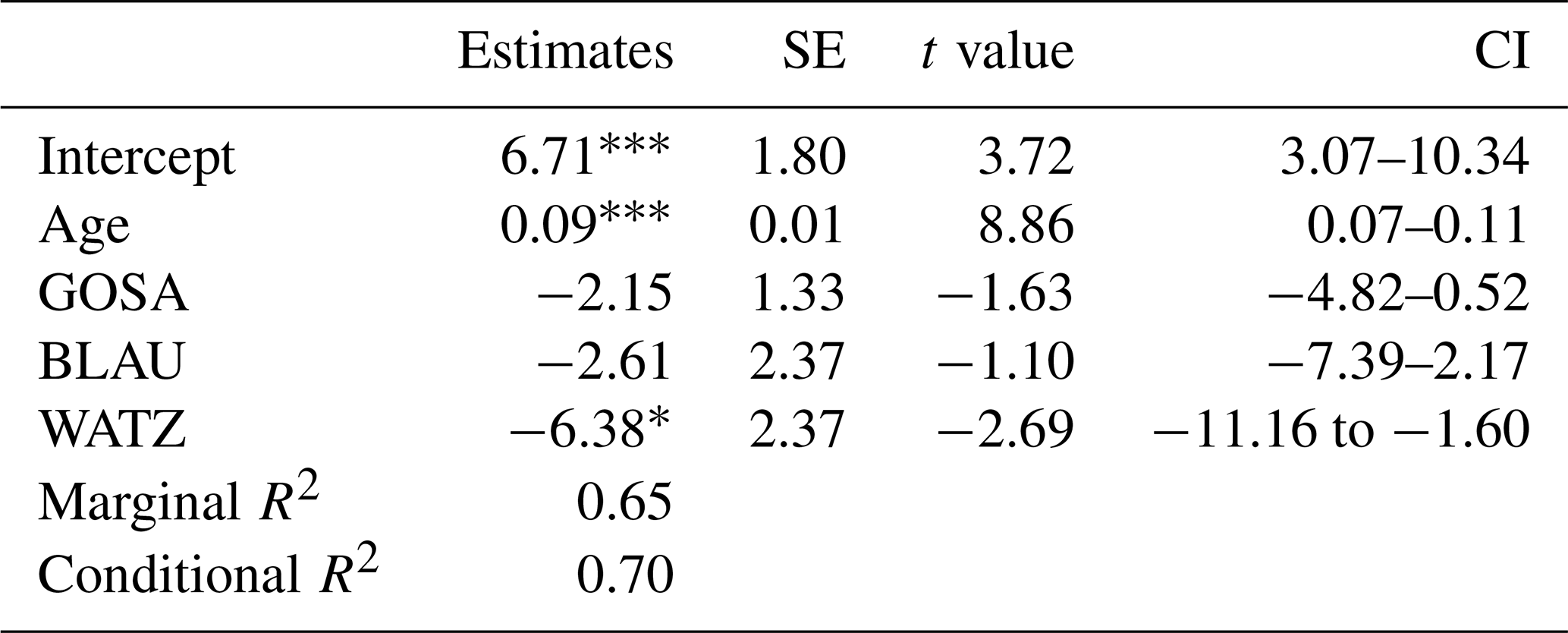
With an increase in age (time since deglaciation), total plant frequency per plot increased (Fig. 5a, Table 3) at a decelerating speed, with the lowest frequency for the youngest plots (ca. 10 years after deglaciation; median = 11.5), followed by plots with ca. 40 years since deglaciation (72.5), ca. 70 years (97.0), and 100–150 years (98.5). Species cover per plot, though, increased almost linearly (Fig. 5b, Table 4); still, it was hardly exceeding 10 % during the first 70 years since deglaciation and not more than 15 % after 150 years. In both cases, no significant interaction and no significant glacier effect was detected.
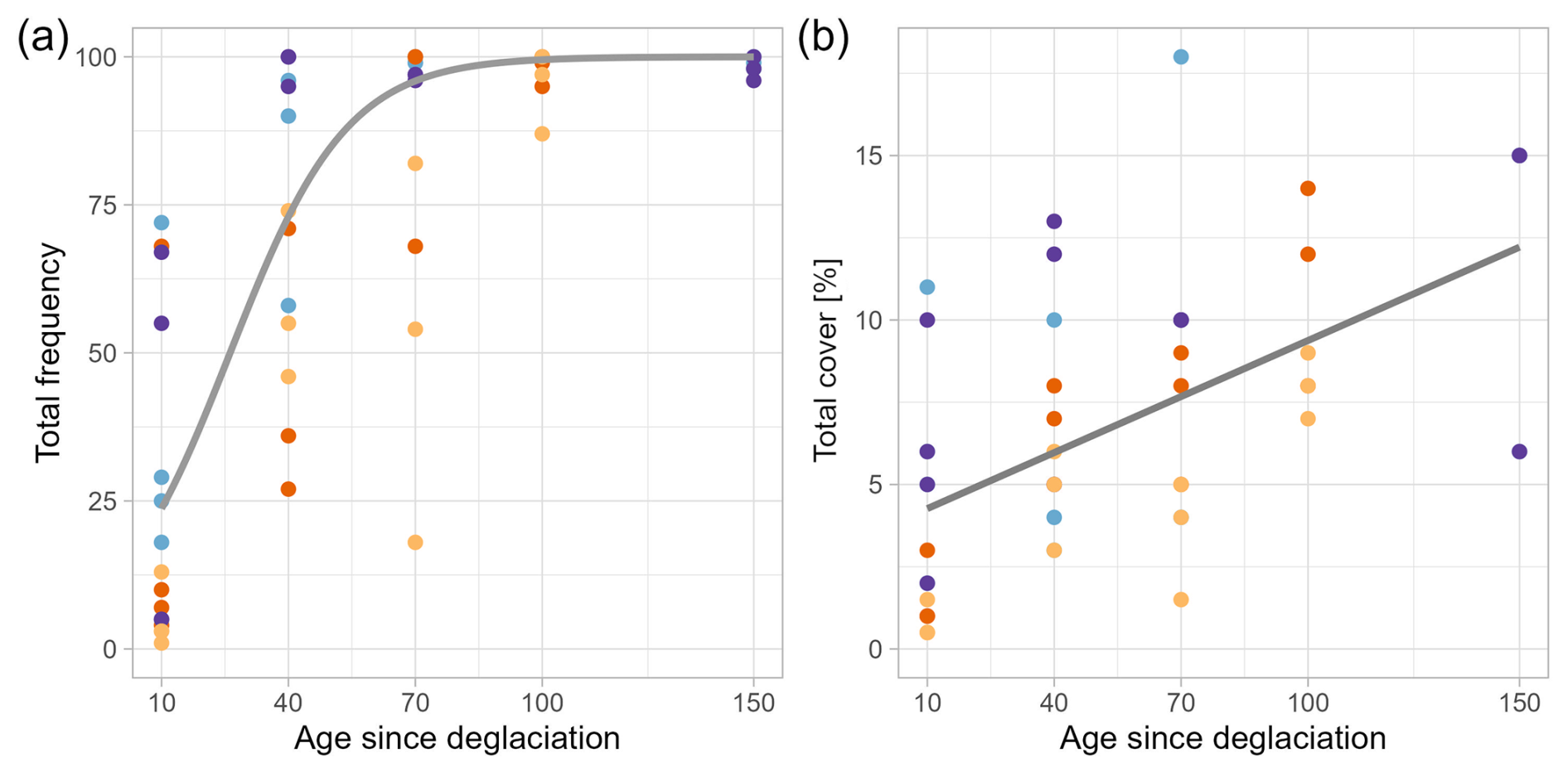
Figure 5Total frequency (a) and total cover of vascular plants (b) per plot of the four glacier forefields in relation to age since deglaciation. Please note that frequency data refer to 2017 (Dachstein glaciers) and 2018 (Berchtesgaden glaciers), while cover of vascular plants refers to 2022.
Table 3Summary statistics relating plant frequency to age since deglaciation with glacier forefield nested in a mountain range as random factor. A generalized mixed effects model with binomial error was used (corresponding to Fig. 5a) (OR: odds ratio, SE: standard error, CI: confidence interval). * 0.01 < p≤0.05; 0.001 <p≤0.01.
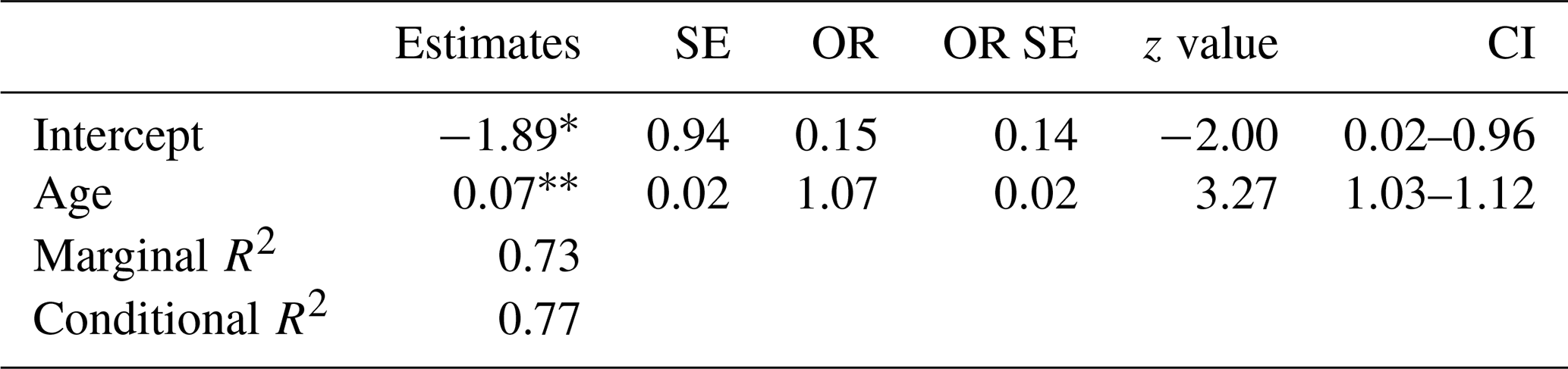
Table 4Summary statistics relating vascular plant cover (data from 2022) to age since deglaciation with glacier and glacier forefield nested in a mountain range as random factor. A mixed effects model was used (corresponding to Fig. 5b) (SE: standard error, CI: confidence interval). * 0.01 <p≤0.05; p<0.001.
In our study system, no saturation in the increase of species richness was discernible, so far, and the total plant cover remained rather low. This is much less than what is known from Central Alps glacier forefields that are by far better studied and in which a saturation in the increase of plant species richness was discernible after 60–100 years and where after 150 years much higher proportions of vegetation cover were reported (e.g. Burga, 1999; Nagl and Erschbamer, 2010; Eichel, 2019; Eichel et al., 2023; see below). It is therefore remarkable that, in comparison to the development in species richness and species cover, the speed of the succession process in the glacier forefields of the Northern Limestone Alps is much slower. This is even more striking, since the analyses using total species cover rather than species frequency per plot started even 4 to 5 years later; therefore, the time for succession was longer. These general patterns, i.e. increase of species richness and cover with age, have also been observed in the other glacier forefields (overviews in Cantera et al. 2024; Cauvy-Fraunié and Dangles, 2019; Ficetola et al., 2021). At the Rotmoosferner forefield (Austria), vegetation cover reached ca. 50 % within ca. 40 years and ca. 70 % within ca. 110 years since deglaciation (Nagl and Erschbamer, 2010). At the Morteratsch glacier forefield (Switzerland), individual species reached cover classes > 75 % in less than 30 years (Burga, 1999). Similarly, the studies by Eichel (2019) and Eichel et al. (2023) for the Turtmann Glacier (Switzerland) indicate considerable vegetation cover on forefields that were older than 70 years. Species richness changes during succession are inherently difficult to compare, because species richness largely depends on plot sizes. Also using 1 m2 plots, species richness reached almost 20 species within 40 years in the Rotmoos Glacier forefield (Nagl and Erschbamer, 2010; Erschbamer and Caccianiga, 2016). This is comparable to the Hallstätter Glacier, but it took at least 100 years in the other glacier forefields. Given that the central European species pool consists of many more calciphilous than acidophilous species (Ewald, 2003), this is even more striking.
We hypothesize three main processes that might have contributed to the differences in species richness and colonization speed between our study sites and other studied Alpine glacier forefields, all related to the geological and geomorphological setting of the different glaciers.
Firstly, the physical and chemical properties of the substrate (not speaking of soil, yet) is different from gneiss and granite, mainly found in the Central Alps. The geological situation in the Ötztal Alps is even more complex, with considerable influences by mica schists and marble, leading to the colonization of calciphile species (Frank et al., 1987). This heterogeneity adds to the complexity in comparison to the almost exclusively karstic limestone bedrock of the Northern Limestone Alps sites. Still, vegetation composition between limestone (calcareous) and gneiss (siliceous) substrates is remarkably different (please consider, however, the heterogeneity of the substrate mentioned above).
Secondly, the karstic situation results in quick drainage of water into the ground. This slows down the soil accumulation processes, as developing particles and accumulating nutrients get easily washed away (Bauer and Plan, 2022). Also, this leads to rather dry substrates during the summer months, which might have a stronger effect on plant species colonization, composition, and growth compared to other substrate properties (Michalet et al., 2002).
Thirdly, many of the previously studied Central Alps glaciers are valley glaciers with hillslopes above them that were free of ice for many hundreds of years and covered by species-rich vegetation. When these glaciers retreated, the forefields could easily be colonized by propagule dispersal from above, i.e. seeds or vegetative material falling mainly downwards, following gravity and the course of the running water (Raffl et al., 2006b), which is a rather fast process. In contrast, on the Dachstein Massif and the Berchtesgaden Alps, glaciers are generally wider and only few ice-free hillslopes exist, with mostly sparse rocky and scree vegetation above the glacier forefields. Colonizing plants hence must have largely moved upwards, which is a much slower process. Additionally, disturbances can play a major role in setting back vegetation succession (Wojcik et al., 2021). Having observed the sites for more than 8 years, this might be a reason, especially at the Blaueis and to a lesser degree at the Watzmann glacier, but it seems to be less likely at the two Dachstein glacier monitoring plots. Therefore (i) propagule pressure might have been much lower and (ii) most of the species observed in the glacier forefields needed to move upwards (from denser vegetation below) rather than falling downwards, which is a much slower process. In conclusion, we hypothesize a largely reduced colonization rate of the succession process in the glacier forefields of the Northern Limestone Alps compared to the Central Alps and Southern Alps.
The intention of this paper is to introduce the BDGF platform and provide initial baseline data for the beginning of a comprehensive study of glacier forefield successions in carbonate landscapes. It is clear that the study of vegetation succession of glacier forefields needs to be complemented by functional approaches in addition to simple observations of changes in species richness and cover (see Erschbamer and Mayer, 2011; Fickert et al., 2017; Losapio et al., 2025), because changes in abiotic and biotic conditions can be expected which are likely to be reflected in the functional characterization of communities (see also McGill et al., 2006). Some initial results have been obtained by Boehm (2023), studying how a suite of functional traits changed with age since deglaciation, using traits from the TRY database (Kattge et al., 2020). Measuring leaf traits in situ, Fels (2022) observed stronger patterns of decline in SLA (specific leaf area, the ratio of leaf area to leaf dry mass) with age since deglaciation. Therefore, we aim to make use of locally measured traits based on Rosbakh et al. (2022), while complementing missing values by on-site sampling and subsequent trait measurements.
To go beyond the chronosequence approach, we plan to validate the obtained results (e.g. the observed increase in plant richness over time) through a palaeoecological approach, leveraging lake sediments from glacier forefield lakes as environmental archives. To this end, we collected lake sediments from Oberer Eissee (an ice lake originating from the 1980s) below the Hallstätter Glacier and analysed the sedimentary ancient DNA pool (similar to Liu et al., 2021, 2024). Palaeogenomic analyses, such as shotgun sequencing of the entire DNA pool, allow for the investigation of succession at an ecosystem scale (Liu et al., 2024). By expanding such analyses to the Unterer Eissee, originating from the late 19th century (or even expanding to other lakes beneath already studies forefields), one can get a broader picture of successional development, covering a larger taxonomic suite of species as well as a larger area by maintaining the sequential (temporal) development of the vegetation in the annual strata of the lake sediments.
As glacier forefield bacterial communities also underlie succession (Hämmerli et al., 2007; Junker et al., 2021; Zumsteg et al., 2012), expanding the ecological surveys to rhizosphere microbiome assemblies could yield novel insights into the functional development of the interactions in the plant–soil–microbiome composition. The prospect of this approach has recently been shown in an initial study based on the rhizosphere microbiome assembly of three plant species along the chronosequence of the Hallstätter Glacier (Wicaksono et al., 2024). Similarly, the microbiome development can be extracted from lake cores of the Oberer Eissee and Unterer Eissee.
Additionally, new insights may be obtained by linking both macrobiome and microbiome succession to the successive weathering and erosion of glacier forefields. Although weathering processes and sediment production are generally slow, the presence of biota can significantly modify chemical reactions and thus instigate feedback processes that affect reaction rates and substrate conditions. Because particulate residues of carbonate weathering are generally limited, the influx of aeolian dust could also play a much greater role compared to glacier forefields developed on silicate bedrock, and this provides another avenue of research at the BDGF platform.
The microtopography of glacier forefields also results in heterogeneous microclimatic conditions in glacier forelands, which might result in microclimatic changes much larger than those of decades of observed climate warming (Marta et al., 2023). These microtopographic conditions are hypothesized to differ from macroclimatic conditions and mediate the impacts on vegetation development (Scherrer and Körner, 2011; Lembrecht et al., 2018). Therefore, a survey of the microclimate with soil temperature loggers installed in 2024 at each plot to record temporal changes in subsurface temperature can provide insights into microclimatic conditions and their influence on ecological processes.
Code and data are available at https://doi.org/10.5281/zenodo.17064649 (Hecht and Kühn, 2025).
IK conceived the idea, established the BDGF platform, performed fieldwork, assisted with the analyses, and wrote the first draft, CH assisted with the fieldwork, performed the analyses, and contributed to writing. UH and DS outlined planned studies in the future, discussed the text, and contributed to text writing. All authors contributed to revising the text.
The contact author has declared that none of the authors has any competing interests.
Publisher’s note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. While Copernicus Publications makes every effort to include appropriate place names, the final responsibility lies with the authors.
GIS layers for glacier extent in the Dachstein massif were provided by Klaus Reingruber (Blue Sky Wetteranalysen, Attnang, Austria). We would like to thank the staff of Blaueishütte, Adamekhütte, and Simonyhütte for their generous support. Further, we are thankful to our enduring field assistants Ken Uhlig, Reed Anderson, Leona Böhm, Angelo Gioia, Ramesh Glückler, Izabella Baisheva, Rahel Paasch, Hannes Petersen, and Jonas Büchner. The Berchtesgaden National Park provided indispensable logistics as well as administrative support. All additional necessary permits were provided by Österreichische Bundesforste, Regierung von Oberösterreich (Abteilung Naturschutz), Regierung von Oberbayern (Sachgebiet Naturschutz), and Landratsamt Berchtesgaden.
Brigitta Erschbamer, Hermann Heilmeier, and an anonymous reviewer provided valuable comments to improve the paper.
This research has been supported by the Programme-Oriented Funding (POF) of the Federal Ministry for Education and Research to the Helmholtz Association.
This paper was edited by Hermann Heilmeier and reviewed by Brigitta Erschbamer and one anonymous referee.
Aeschimann, D., Lauber, K., Moser, D. M., and Theurillat, J. P.: Flora alpina, Haupt, Bern, 334 pp., ISBN 978-3-258-06600-4, 2004.
Bauer, C. and Plan, L.: Karst Landscapes in Austria, in: Landscapes and Landforms of Austria, edited by: Embleton-Hamann, C., Springer International Publishing, Cham, 87–104, https://doi.org/10.1007/978-3-030-92815-5_5, 2022.
Boehm, L.: Functional plant traits and primary succession on chronosequences in glacier forefields of the Northern Limestone Alps, MS thesis, University of Leipzig, Faculty of Life Sciences Institute of Biology, Leipzig, Germany, 2023.
Bosson, J. B., Huss, M., Cauvy-Fraunié, S., Clément, J. C., Costes, G., Fischer, M., Poulenard, J., and Arthaud, F.: Future emergence of new ecosystems caused by glacial retreat, Nature, 620, 562–569, https://doi.org/10.1038/s41586-023-06302-2, 2023.
Burga, C. A.: Vegetation development on the glacier forefield Morteratsch (Switzerland), Appl. Veg. Sci., 2, 17–24, https://doi.org/10.2307/1478877, 1999.
Cannone, N. and Piccinelli, S.: Changes of rock glacier vegetation in 25 years of climate warming in the Italian Alps, Catena, 206, https://doi.org/10.1016/j.catena.2021.105562, 2021.
Cannone, N., Diolaiuti, G., Guglielmin, M., and Smiraglia, C.: Accelerating climate change impacts on alpine glacier forefield ecosystems in the European Alps, Ecol. Appl., 18, 637–648, https://doi.org/10.1890/07-1188.1, 2008.
Cantera, I., Carteron, A., Guerrieri, A., Marta, S., Bonin, A., Ambrosini, R., Anthelme, F., Azzoni, R. S., Almond, P., Alviz Gazitúa, P., Cauvy-Fraunié, S., Ceballos Lievano, J. L., Chand, P., Chand Sharma, M., Clague, J., Cochachín Rapre, J. A., Compostella, C., Cruz Encarnación, R., Dangles, O., Eger, A., Erokhin, S., Franzetti, A., Gielly, L., Gili, F., Gobbi, M., Hågvar, S., Khedim, N., Meneses, R. I., Peyre, G., Pittino, F., Rabatel, A., Urseitova, N., Yang, Y., Zaginaev, V., Zerboni, A., Zimmer, A., Taberlet, P., Diolaiuti, G. A., Poulenard, J., Thuiller, W., Caccianiga, M., and Ficetola, G. F.: The importance of species addition “versus” replacement varies over succession in plant communities after glacier retreat, Nat. Plants, 1–12, https://doi.org/10.1038/s41477-023-01609-4, 2024.
Carvalho, J. C., Cardoso, P., and Gomes, P.: Determining the relative roles of species replacement and species richness differences in generating beta-diversity patterns, Global Ecol. Biogeogr., 21, 760–771, https://doi.org/10.1111/j.1466-8238.2011.00694.x, 2012.
Cauvy-Fraunié, S. and Dangles, O.: A global synthesis of biodiversity responses to glacier retreat, Nature Ecology & Evolution, 3, 1675–1685, https://doi.org/10.1038/s41559-019-1042-8, 2019.
CLARISIA: Climate-Air-Information-System for Upper Austria, https://www.doris.at/themen/umwelt/clairisa.aspx (last access: 3 September 2025), 2015.
Coaz, J.: Erste Ansiedlung phanerogamer Pflanzen auf von Gletschern verlassenem Boden, in: Mittheilungen der Naturforschenden Gesellschaft in Bern, vol. Nr. 1169-1194 (1887), Bern, 3–12, https://www.biodiversitylibrary.org/item/41949 (last access: 4 September 2025), 1887.
Crawley, M. J.: The R Book, 2nd ed., Wiley, 1076 pp., ISBN 0470973927, 2012.
Eichel, J.: Vegetation Succession and Biogeomorphic Interactions in Glacier Forelands, in: Geomorphology of Proglacial Systems, edited by: Heckmann, T. and Morche, D., 327–349, ISBN 978-3-319-94182-0, 2019.
Eichel, J., Draebing, D., Winkler, S., and Meyer, N.: Similar vegetation-geomorphic disturbance feedbacks shape unstable glacier forelands across mountain regions, Ecosphere, 14, e4404, https://doi.org/10.1002/ecs2.4404, 2023.
Erschbamer, B. and Caccianiga, M. S.: Glacier Forelands: Lessons of Plant Population and Community Development, in: Progress in Botany, vol. 78, edited by: Cánovas, F. M., Lüttge, U., and Matyssek, R., Springer, Berlin, Heidelberg, 259–284, https://doi.org/10.1007/124_2016_4, 2016.
Erschbamer, B. and Mayer, R.: Can successional species groups be discriminated based on their life history traits? A study from a glacier foreland in the Central Alps, Plant Ecol. Divers., 4, 341–351, https://doi.org/10.1080/17550874.2012.664573, 2011.
Erschbamer, B., Bitterlich, W., and Raffl, C.: Die Vegetation als Indikator für die Bodenbildung im Gletschervorfeld des Rotmoosferners (Obergurgl, Ötztal, Nordtirol), Berichte des naturwissenschaftlich-medizinischen Vereins in Innsbruck, 86, 107–122, 1999.
Erschbamer, B., Niederfriniger Schlag, R., Carnicero, P., and Kaufmann, R.: Long-term monitoring confirms limitations of recruitment and facilitation and reveals unexpected changes of the successional pathways in a glacier foreland of the Central Austrian Alps, Plant Ecol., 224, 373–386, https://doi.org/10.1007/s11258-023-01308-2, 2023.
Ewald, J.: The calcareous riddle: Why are there so many calciphilous species in the Central European flora?, Folia Geobot., 38, 357–366, https://doi.org/10.1007/BF02803244, 2003.
Fels, L.: Changes in plant community composition along a successional gradient in the forefields of the Blaueis, Northern Limestone Alps, MS thesis, University of Vienna, Vienna, Austria, https://utheses.univie.ac.at/detail/65542# (last access: 4 September 2025), 2022.
Ficetola, G. F., Marta, S., Guerrieri, A., Gobbi, M., Ambrosini, R., Fontaneto, D., Zerboni, A., Poulenard, J., Caccianiga, M., and Thuiller, W.: Dynamics of ecological communities following current retreat of glaciers, Annual Rev. Ecol. Evol. Syst., 52, 405–426, https://doi.org/10.1146/annurev-ecolsys-010521-040017, 2021.
Fickert, T., Grüninger, F., and Damm, B.: Klebelsberg revisited: did primary succession of plants in glacier forelands a century ago differ from today?, Alpine Bot., 127, 17–29, https://doi.org/10.1007/s00035-016-0179-1, 2017.
Frank, W., Hoinkes, G., Purtscheller, F., and Thöni, M.: The Austroalpine unit west of the Hohe Tauern: the Ötztal-Stubai complex as an example for the Eo-Alpine metamorphic evolution, in: Geodynamics of the Eastern Alps, edited by: Flügel, H. W. and Faupl, P., Franz Deuticke, Vienna (Austria), 179–225, ISBN 978-3-7005-4528-6, 1987.
Frenzel, B: Exkursionsberichte der deutsch-österreichischen Gletscherkurse von 1951 und 1957 ins Gebiet des Schwarzensteinkees incl. Karte der Pflanzengesellschaften des Vorfelds von 1957 (manuscripts), 1951/1957.
Ganss, O. and Grünfelder, S.: Geologie der Berchtesgadener und Reichenhaller Alpen: Eine Einführung in die Gesteinsbildung, Gebirgsbildung und Landschaftsgeschichte, Plenk, Berchtesgaden, ISBN 3922590101, 1979.
Geoland.at: Open Data of Digital Elevation Model were downloaded from OpenDEMEurope: Produced using Copernicus data and information funded by the European Union, Geoland.at (2019), GEOPORTALE NAZIONALE - AccessConstraints: Nessuno – None, https://opendem.info (last access: 3 September 2025), 2019.
Grims, F.: Über die Besiedlung der Vorfelder einiger Dachsteingletscher (Oberösterreich), Stapfia, 10, 203–233, 1982.
Hagg, W.: Digitale Aufbereitung historischer Gletscherkarten in Bayern, Mitteilungen der Geographischen Gesellschaft in München, 88, 67–78, 2006.
Hämmerli, A., Waldhuber, S., Miniaci, C., Zeyer, J., and Bunge, M.: Local expansion and selection of soil bacteria in a glacier forefield, Eur. J. Soil Sci., 58, 1437–1445, https://doi.org/10.1111/j.1365-2389.2007.00948.x, 2007.
Hanusch, M., He, X., Ruiz-Hernandez, V., and Junker, R. R.: Succession comprises a sequence of threshold-induced community assembly processes towards multidiversity, Communications Biology, 5, 424, https://doi.org/10.1038/s42003-022-03372-2, 2022.
Hecht, C. and Kühn, I.: Dataset for: “Introducing a Glacier Forefield Monitoring Site Network to Understand Succession in the Northern Limestone Alps”, Zenodo [code and data set], https://doi.org/10.5281/zenodo.17064649, 2025.
Helfricht, K.: Veränderungen des Massenhaushaltes am Hallstätter Gletscher seit 1856, Diploma thesis, Institut für Meteorologie und Geophysik, Leopold-Franzens-Universität Innsbruck, Innsbruck, Austria, https://epic.awi.de/id/eprint/32204/ (last access: 4 September 2025), 2009.
Jochimsen, M.: Vegetationsentwicklung im hochalpinen Neuland. (Beobachtungen an Dauerflächen im Gletschervorfeld 1958-1962), Berichte des naturwissenschaftlichen-medizinischen Verein Innsbruck, 53, 109–123, 1963.
Junker, R. R., He, X., Otto, J. C., Ruiz-Hernandez, V., and Hanusch, M.: Divergent assembly processes? A comparison of the plant and soil microbiome with plant communities in a glacier forefield, FEMS Microbiol. Ecol., 97, https://doi.org/10.1093/femsec/fiab135, 2021.
Kattge, J., Bonisch, G., Diaz, S., Lavorel, S., Prentice, I. C., Leadley, P., Tautenhahn, S., Werner, G. D. A., Aakala, T., Abedi, M., Acosta, A. T. R., Adamidis, G. C., Adamson, K., Aiba, M., Albert, C. H., Alcantara, J. M., Alcazar, C. C., Aleixo, I., Ali, H., Amiaud, B., Ammer, C., Amoroso, M. M., Anand, M., Anderson, C., Anten, N., Antos, J., Apgaua, D. M. G., Ashman, T. L., Asmara, D. H., Asner, G. P., Aspinwall, M., Atkin, O., Aubin, I., Baastrup-Spohr, L., Bahalkeh, K., Bahn, M., Baker, T., Baker, W. J., Bakker, J. P., Baldocchi, D., Baltzer, J., Banerjee, A., Baranger, A., Barlow, J., Barneche, D. R., Baruch, Z., Bastianelli, D., Battles, J., Bauerle, W., Bauters, M., Bazzato, E., Beckmann, M., Beeckman, H., Beierkuhnlein, C., Bekker, R., Belfry, G., Belluau, M., Beloiu, M., Benavides, R., Benomar, L., Berdugo-Lattke, M. L., Berenguer, E., Bergamin, R., Bergmann, J., Carlucci, M. B., Berner, L., Bernhardt-Romermann, M., Bigler, C., Bjorkman, A. D., Blackman, C., Blanco, C., Blonder, B., Blumenthal, D., Bocanegra-Gonzalez, K. T., Boeckx, P., Bohlman, S., Bohning-Gaese, K., Boisvert-Marsh, L., Bond, W., Bond-Lamberty, B., Boom, A., Boonman, C. C. F., Bordin, K., Boughton, E. H., Boukili, V., Bowman, D., Bravo, S., Brendel, M. R., Broadley, M. R., Brown, K. A., Bruelheide, H., Brumnich, F., Bruun, H. H., Bruy, D., Buchanan, S. W., Bucher, S. F., Buchmann, N., Buitenwerf, R., Bunker, D. E., et al.: TRY plant trait database – enhanced coverage and open access, Glob. Change Biol., 26, 119–188, https://doi.org/10.1111/gcb.14904, 2020.
Krause, H. G. and Frenzel, B.: Die Entwicklung der Vegetation eines zentralalpinen Gletschervorfeldes seit dem Jahr 1957, Jahrbuch zum Schutz der Bergwelt, 62, 189–226, 1997.
Kuhn, M., Schlosser, E., and Span, N.: Eastern Alpine glacier activity and climatic records since 1860, Ann. Glaciol., 24, 164–168, https://doi.org/10.3189/S0260305500012118, 1997.
Lembrechts, J. J., Lenoir, J., Nuñez, M. A., Pauchard, A., Geron, C., Bussé, G., Milbau, A., and Nijs, I.: Microclimate variability in alpine ecosystems as stepping stones for non-native plant establishment above their current elevational limit, Ecography, 41, 900–909, https://doi.org/10.1111/ecog.03263, 2018.
Liu, S., Kruse, S., Scherler, D., Ree, R. H., Zimmermann, H. H., Stoof-Leichsenring, K. R., Epp, L. S., Mischke, S., and Herzschuh, U.: Sedimentary ancient DNA reveals a threat of warming-induced alpine habitat loss to Tibetan Plateau plant diversity, Nat. Commun., 12, 2995, https://doi.org/10.1038/s41467-021-22986-4, 2021.
Liu, S., Stoof-Leichsenring, K. R., Harms, L., Schulte, L., Mischke, S., Kruse, S., Zhang, C., and Herzschuh, U.: Tibetan terrestrial and aquatic ecosystems collapsed with cryosphere loss inferred from sedimentary ancient metagenomics, Science Advances, 10, eadn8490, https://doi.org/10.1126/sciadv.adn8490, 2024.
Losapio, G., Lee, J. R., Fraser, C. I., Gillespie, M. A. K., Kerr, N. R., Zawierucha, K., Hamilton, T. L., Hotaling, S., Kaufmann, R., Kim, O.-S., Klopsch, C., Liu, Y., Lukashanets, D., Robinson, S. A., and Brown, L. E.: Impacts of deglaciation on biodiversity and ecosystem function, Nature Reviews Biodiversity, 1–15, https://doi.org/10.1038/s44358-025-00049-6, 2025.
Marta, S., Zimmer, A., Caccianiga, M., Gobbi, M., Ambrosini, R., Azzoni, R. S., Gili, F., Pittino, F., Thuiller, W., Provenzale, A., and Ficetola, G. F.: Heterogeneous changes of soil microclimate in high mountains and glacier forelands, Nat. Commun., 14, 5306, https://doi.org/10.1038/s41467-023-41063-6, 2023.
Mayer, C., Weber, M., Wendt, A., and Hagg, W.: Die bayerischen Gletscher, die verbliebenen Eisreserven Deutschlands, Polarforschung, 89, 1–7, https://doi.org/10.5194/polf-89-1-2021, 2021.
Mayer, D. C.: Das „Ewige Eis auf dem Rückzug“, Akademie aktuell, Zeitschrift der Bayerischen Akademie der Wissenschaften, 4, 30–33, https://badw.de/fileadmin/pub/akademieAktuell/2012/43/0412_11_mayer.pdf (last access: 4 September 2025), 2012.
McGill, B. J., Enquist, B. J., Weiher, E., and Westoby, M.: Rebuilding community ecology from functional traits, Trends Ecol. Evol., 21, 178–185, https://doi.org/10.1016/j.tree.2006.02.002, 2006.
Michalet, R., Gandoy, C., Joud, D., Pagès, J.-P., and Choler, P.: Plant community composition and biomass on calcareous and siliceous substrates in the Northern French Alps: comparative effects of soil chemistry and water status, Arct. Antarct. Alp. Res., 34, 102–113, https://doi.org/10.1080/15230430.2002.12003474, 2002.
Moser, R.: Dachsteingletscher und deren Spuren im Vorfeld, Musealverein Hallstatt, Hallstatt (Austria), https://www.zobodat.at/pdf/GmuGeoStud_2_0237-0246.pdf (last access: 4 September 2025), 1997.
Moser, R. and Vareschi, V.: Die Pflanzen der Moränen des Dachsteins, Jahrbuch des Oberösterreichischen Musealvereines, 104, 181–200, 1959.
Nagl, F. and Erschbamer, B.: Pflanzliche Sukzession im Gletschervorfeld: Vegetation und Besiedelungsstrategien, Publikationen Alpine Forschungsstelle Obergurgl, 1, 121–143, 2010.
Nicklas, L., Walde, J., Wipf, S., Lamprecht, A., Mallaun, M., Rixen, C., Steinbauer, K., Theurillat, J.-P., Unterluggauer, P., Vittoz, P., Moser, D., Gattringer, A., Wessely, J., and Erschbamer, B.: Climate change affects vegetation differently on siliceous and calcareous summits of the European Alps, Front. Ecol. Evol., 9, https://doi.org/10.3389/fevo.2021.642309, 2021.
Nicolussi, K., Le Roy, M., Schlüchter, C., Stoffel, M., and Wacker, L.: The glacier advance at the onset of the Little Ice Age in the Alps: New evidence from Mont Miné and Morteratsch glaciers, Holocene, 32, 624–638, https://doi.org/10.1177/09596836221088247, 2022.
Pauli, H., Gottfried, M., Lamprecht, A., Niessner, S., Rumpf, S., and Winkler, M.: The GLORIA field manual – standard multi-summit approach, supplementary methods and extra approaches, GLORIA-Coordination, Austrian Academy of Sciences & University of Natural Resources and Life Sciences, Vienna, https://www.gloria.ac.at/methods/manual (last access: 4 September 2025), 2015.
R Core Team: R: A language and environment for statistical computing, R Foundation for Statistical Computing, Vienna, Austria, https://cran.r-project.org/doc/manuals/r-release/fullrefman.pdf (last access: 4 September 2025), 2023.
Raffl, C., Mallaun, M., Mayer, R., and Erschbamer, B.: Vegetation succession pattern and diversity changes in a glacier valley, Central Alps, Austria, Arct. Antarct. Alp. Res., 38, 421–428, https://doi.org/10.1657/1523-0430(2006)38[421:VSPADC]2.0.CO;2, 2006a.
Raffl, C., Schönswetter, P., and Erschbamer, B.: “Sax-sess” – genetics of primary succession in a pioneer species on two parallel glacier forelands, Mol. Ecol., 15, 2433–2440, https://doi.org/10.1111/j.1365-294X.2006.02964.x, 2006b.
Rosbakh, S., Chalmandrier, L., Phartyal, S., and Poschlod, P.: Inferring community assembly processes from functional seed trait variation along elevation gradient, J. Ecol., https://doi.org/10.1111/1365-2745.13955, 2022.
Scheidleder, A., Boroviczeny, F., Graf, W., Hofmann, T., Mandl, G. W., Schubert, G., Stichler, W., Trimborn, P., and Kralik, M.: Pilotprojekt „Karstwasser Dachstein “, Band 2: Karsthydrologie und Kontaminationsrisiko von Quellen, Geologische Bundesanstalt & Umweltbundesamt, Vienna, https://www.zobodat.at/publikation_volumes.php?id=34258 (last access: 4 September 2025), 2001.
Scherrer, D. and Körner, C.: Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming, J. Biogeogr., 38, 406–416, https://doi.org/10.1111/j.1365-2699.2010.02407.x, 2011.
Wicaksono, W. A., Mora, M., Bickel, S., Berg, C., Kühn, I., Cernava, T., and Berg, G.: Rhizosphere assembly alters along a chronosequence in the Hallstätter glacier forefield (Dachstein, Austria), FEMS Microbiol. Ecol., fiae005, https://doi.org/10.1093/femsec/fiae005, 2024.
Wojcik, R., Eichel, J., Bradley, J. A., and Benning, L. G.: How allogenic factors affect succession in glacier forefields, Earth-Sci. Rev., 218, https://doi.org/10.1016/j.earscirev.2021.103642, 2021.
Zumsteg, A., Luster, J., Göransson, H., Smittenberg, R. H., Brunner, I., Bernasconi, S. M., Zeyer, J., and Frey, B.: Bacterial, archaeal and fungal succession in the forefield of a receding glacier, Microb. Ecol., 63, 552–564, https://doi.org/10.1007/s00248-011-9991-8, 2012.