the Creative Commons Attribution 4.0 License.
the Creative Commons Attribution 4.0 License.
 | 08 Jul 2026
| 08 Jul 2026
Landscape context outweighs field-scale features in structuring bird communities in West African smallholder rice agroecosystems
Paula Lopes
Inês Lacerda
Patrícia Chaves
Ricardo F. de Lima
Mutaro Camará
Miguel Lecoq
Jorge M. Palmeirim
Ana Rainho
Understanding how local management and landscape structure influence biodiversity in tropical croplands is crucial for reconciling food production and conservation. Here, we examined how field- and landscape-scale characteristics relate to bird species richness in lowland rain-fed rice fields of northern Guinea-Bissau. Birds were surveyed along nine transects repeatedly sampled during two rice-growing cycles (2021–2022), and species were assigned to four trophic guilds: frugivores, granivores, invertivores, and omnivores. Field variables (parcel area, tree cover, field width) and landscape metrics (fragmentation, diversity, edge complexity) were derived from satellite imagery across multiple spatial scales. Species richness for each transect survey was estimated using Bayesian models, and linear mixed models were used to relate richness to environmental predictors and to principal ecological gradients summarizing field- and landscape-level structure.
A total of 150 bird species were identified, with an estimated mean species richness of 18.2 species per transect survey. At the field scale, tree cover increased invertivore and frugivore richness by 17.0 % and 16.4 % of the standard deviation, respectively, while wider fields supported higher overall richness and higher richness of granivores and omnivores, with increases of 2.9 %, 5.6 %, 18.4 %, and 14.0 % of the standard deviation, respectively. At the landscape level, bird richness varied with composition and configuration in a scale- and guild-dependent manner: frugivores, invertivores, and omnivores responded primarily at broader scales (1600 m radius around the rice fields), whereas the entire community and granivores showed stronger associations with intermediate-scale attributes (800 m radius). When jointly considering field- and landscape-scale attributes, bird richness showed stronger associations with landscape-scale gradients than with field-scale characteristics. Landscape characteristics significantly influenced the richness of the entire community as well as all trophic guilds considered. In contrast, field-level features affected only frugivores, invertivores, and omnivores, with the richness of these guilds increasing with field openness. These results highlight the context-dependent ecological meaning of commonly used heterogeneity metrics and suggest that sustaining bird richness in smallholder rice systems is likely to benefit from landscape-level planning complemented by field-scale management actions, such as retaining scattered trees.
- Article
(3096 KB) - Full-text XML
-
Supplement
(4688 KB) - BibTeX
- EndNote




More than one-third of the Earth's land surface is currently devoted to agriculture (FAO, 2024; Tubiello et al., 2023), and bird species associated with agroecosystems are experiencing alarming global population declines (Perrings and Halkos, 2015; Rigal et al., 2023; Şekercioglu et al., 2019). Long-term monitoring in temperate regions has revealed steep and persistent reductions: in the European Union, agricultural and grassland species have declined by 35 % since 1980 (Burns et al., 2021), while in North America, farmland birds have lost 74 % of their abundance since 1966 (Stanton et al., 2018). Similar patterns have been observed elsewhere; in Japan, farmland birds showed a marked contraction in geographic range at around the turn of the century (Amano and Yamaura, 2007). Although systematic monitoring in Africa remains limited, an 18-year study in Nigeria found that bird species undergoing declines were predominantly associated with open and grassland habitats (Ishong et al., 2022). Across Africa, tropical grassland biodiversity is generally declining due to the ongoing degradation and transformation of these ecosystems, with some regions having lost over 30 % of native grassland cover in recent decades (Newbold et al., 2017; Vukeya et al., 2025).
Rice, as both a cornerstone of food security and a dominant land use in tropical regions (Global Rice Science Partnership, 2013), exemplifies the challenge of reconciling agricultural production with biodiversity conservation. In 2023, Africa had over 18.7 million ha under rice cultivation, most of it in West Africa (Jiang et al., 2025), where production is largely carried out by smallholders using labour-intensive practices and minimal chemical inputs (Temudo, 2011). Although such systems often produce lower yields than mechanized agriculture, they are generally associated with higher biodiversity and the maintenance of key ecosystem functions, as reported for Southeast Asia rice systems (Propper et al., 2024). Traditional rice fields can provide habitats for a wide range of bird species, especially waterbirds and open-area specialists (Bos et al., 2006; Cao and Fox, 2009; Fasola and Ruiz, 1996; Weller, 1999; Wymenga and Zwarts, 2010). In some regions, flooded rice paddies act as surrogates for natural wetlands, offering foraging and roosting opportunities (Fasola and Ruiz, 1996). However, the ecological value of rice paddies is highly context dependent and strongly mediated by local management (Tong, 2017).
Rice-growing landscapes are inherently dynamic, and management decisions often vary over time and among farmers in response to social or economic pressures (Temudo, 2011). Differences in landowners' goals, water management, and mechanization generate strong spatial contrasts in habitat structure, influencing key ecological attributes such as vegetation complexity, water availability, and food resources that determine suitability for different bird groups (King et al., 2010). Traditional rice fields in West Africa are often established in rain-fed floodplains and are typically characterized by narrow configurations and irregular edges. Isolated trees or small tree clumps are commonly retained for resource extraction, soil fertility enhancement (e.g. nitrogen fixation), and shade provision, and contribute to soil moisture retention (Garrity et al., 2010). Beyond field-level characteristics, bird diversity in rice agroecosystems is also shaped by the broader landscape context, including land-cover composition, fragmentation, and configuration (Elphick, 2008; Marcolin et al., 2021), with responses often mediated by species' ecological traits (King et al., 2010).
Despite West Africa's importance both as a major rice-producing region and as a centre of high avian diversity, bird communities in its rice fields remain poorly studied (Kouadja et al., 2023; Odoukpe and Yaokokore-Beibro, 2014). Existing research on rice agroecosystems has focused largely on waterbirds, with a strong emphasis on intensive production systems (Bos et al., 2006; Nachuha, 2009; Wymenga and Zwarts, 2010; Yaokororé-Béibro and Odoukpé, 2015; Zwarts et al., 2009); no studies to date have examined how multiple dimensions of landscape structure operate across spatial scales to shape broader bird communities. Moreover, although landscape heterogeneity is often associated with positive biodiversity responses in agricultural systems, similar heterogeneity values may reflect contrasting landscape configurations, in which habitat patches differ in their spatial arrangement and connectivity, leading to different ecological outcomes (Fahrig et al., 2011; Tscharntke et al., 2012). As a result, it remains unclear whether general expectations derived from other regions, such as the positive effects of field- and landscape-scale diversity, apply to low-intensity, smallholder rice landscapes in the tropics. The relative importance of field- versus landscape-scale characteristics in shaping biodiversity responses in these systems also remains poorly understood.
In this study, we provide a functional assessment, based on trophic guilds, of bird species richness in small-scale, rain-fed floodplain rice fields in Guinea-Bissau, based on repeated surveys conducted across two rice-growing cycles. By explicitly separating field-scale structural attributes from landscape-scale gradients of composition and configuration, and by evaluating responses across multiple spatial scales, we aim to examine how bird species richness responds to commonly used measures of landscape heterogeneity in a smallholder rice system, and to compare the relative importance of field- and landscape-scale drivers. Specifically, we address the following questions:
-
How do rice field characteristics, namely parcel area, tree cover, and field width, influence bird species richness at the community level and within trophic guilds?
-
How does landscape composition and configuration, measured by fragmentation, diversity, and edge complexity at multiple spatial scales, influence bird species richness across the surveyed community?
-
How does bird species richness, overall and across trophic guilds, respond to ecological gradients at field and landscape scales, and what is the relative influence of local versus landscape-scale drivers?
We hypothesize that both field- and landscape-scale characteristics would influence bird richness, with responses varying among trophic guilds. At the field scale, we expect that greater structural complexity, reflected by tree and shrub cover, would be associated with higher species richness by increasing the availability of foraging substrates, nesting sites and shelter. We further expect that field width would influence richness by modulating openness and edge availability within rice fields (Longoni, 2010). At the landscape scale, we hypothesize that bird species richness would respond to gradients of composition and configuration across spatial scales, reflecting differences in habitat availability, connectivity, and access to complementary resources (He et al., 2023; Marcolin et al., 2021). We expect that the direction and strength of these relationships would depend on the ecological meaning captured by commonly used heterogeneity metrics, which may vary across landscapes and scales (Berry, 2001; Lee et al., 2024). Finally, we hypothesize that bird richness would be more strongly associated with landscape-scale gradients than with field-scale characteristics, consistent with the influence of the broader spatial context on local communities (Cabral et al., 2021; Redlich et al., 2018).
By linking the multi-scale drivers of bird richness with diet-based functional groupings, our study aims to advance ecological understanding and inform agroecological management and landscape planning, offering guidance for policies that aim to reconcile food production with biodiversity conservation in smallholder rice systems in West Africa.
2.1 Study area
The study was carried out in the Oio region, northern Guinea-Bissau (Fig. 1). The area is flat, with a landscape dominated by savannah woodland, cashew orchards, and rain-fed floodplain rice fields. Due to human activities, particularly fire and shifting agriculture, the prevailing vegetation consists of secondary formations, although some patches of native forest persist (Catarino et al., 2008; dos Reis-Silva et al., 2025). Urban settlements are small, generally housing only a few hundred inhabitants. The climate is typical of tropical savannah, with an average annual temperature of 28 °C and little intra-annual variation. The rainy season occurs from June to November, peaking in August, while the remaining months are dry (The Climate Change Knowledge Portal, 2025).
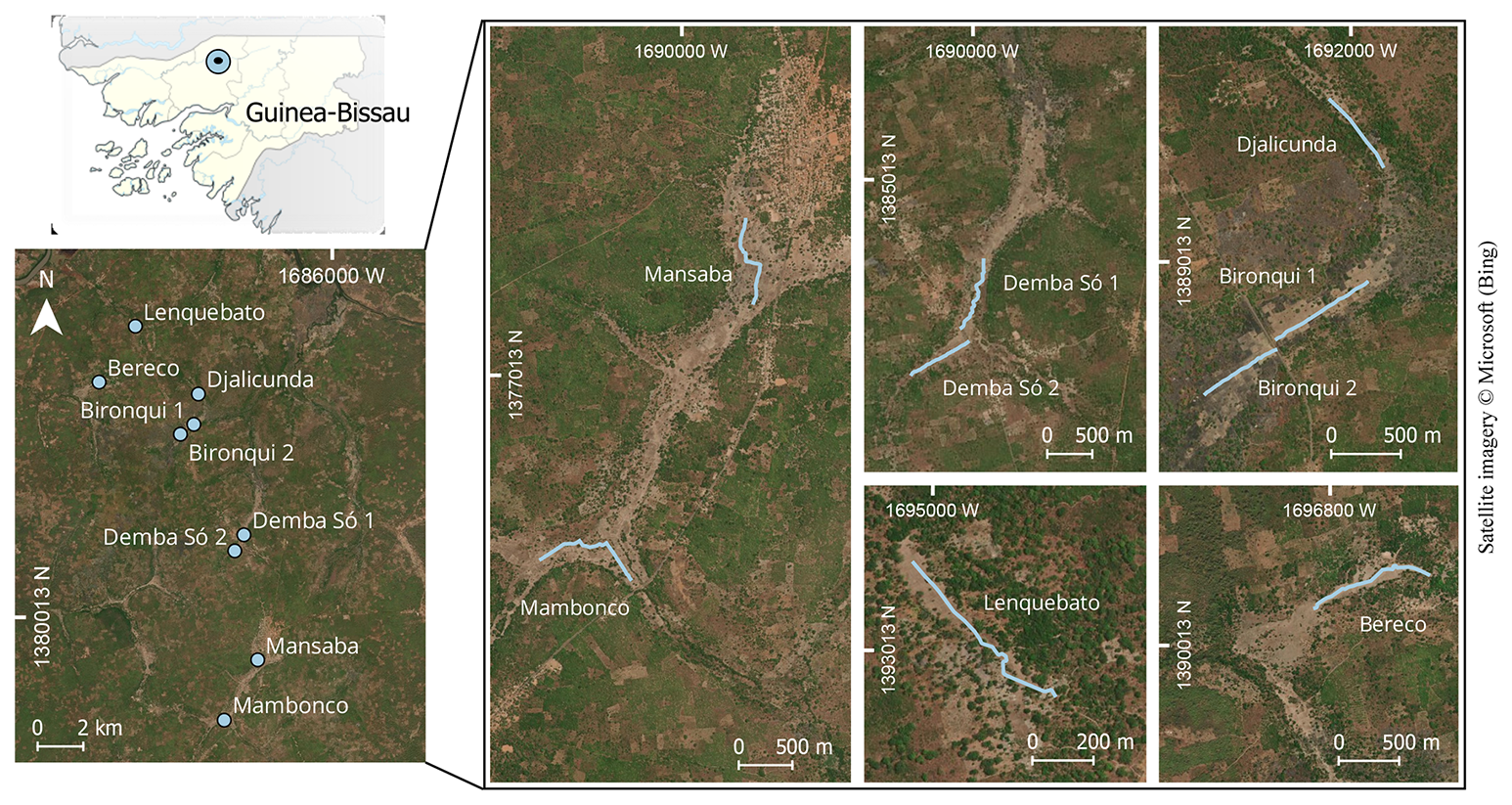
Figure 1Location of the study area in Guinea-Bissau (top left), distribution of the transects across the landscape (bottom left), and location of the nine transects in the floodplain valleys with rice fields (right).
In our study area, rice fields were located along floodplains bordering small watercourses, extending for 1–4 km and varying from a few dozen up to more than 200 m in width. Each field consisted of dozens of parcels separated by earthen bunds – slightly raised earth structures that delineate individual parcels, control water flow, and often support natural vegetation (Fig. S1 in the Supplement). Each parcel has an approximately square shape with an average side length of 50 m. Rice cultivation is carried out almost entirely by women from nearby villages, using manual practices and minimal chemical inputs (Chaves et al., 2024). During the study period, each woman managed her own parcel independently, resulting in a heterogeneous mosaic in which parcels with different rice varieties, management techniques, and growth stages occurred side by side. Sowing began with the onset of the rainy season in late June or July, and harvest took place between December and January. After harvest, fields lay fallow until the next growing cycle, sometimes serving as grazing areas for cattle.
2.2 Bird surveys and trophic guild data
Bird communities were surveyed along nine transects established in seven rice fields and repeatedly visited during the rice-growing cycle. Each transect (610–1200 m in length) crossed dozens of parcels within the same rice field. Transects were surveyed as continuous walks at a constant pace, without fixed stopping points, and all birds seen or heard within the rice fields, from the observer to the field's edge, were recorded (Bibby et al., 2000). In total, 224 transect surveys were conducted. Each survey (transect × date) was treated as an individual sampling unit in the analyses, and richness was not aggregated across repeated visits.
Birds flying over the rice fields were excluded, except for aerial foragers such as swallows and swifts. Surveys began at sunrise and were completed within a 3 h window, during which two transects were surveyed (including travel time), while avoiding adverse weather conditions, such as strong winds or rain. Each transect survey lasted an average of 63 min (standard deviation (SD) = 24 min). All transects were performed by the first author, with some replicates conducted by the second author. Both are experienced field ornithologists familiar with the local avifauna. These main observers were always accompanied by a less experienced assistant who recorded the data and provided additional detections.
Sampling was conducted in three periods: (1) October to December 2021, (2) June to July 2022, and (3) September to December 2022. In 2021, six transects were established across five rice fields near the settlements of Lenquebato, Djalicunda, Bironqui, Bereco, and Demba Só. In 2022, one transect was established in each of three additional rice fields: Demba Só, Mansaba, and Mambonco (Fig. 1). In both years, each transect was surveyed between 10 and 38 times, with a mean of 25 surveys per transect. Sampling effort ranged from 17 surveys in June to 64 in October, averaging 37 per month.
We assembled data on trophic guilds from the AVONET database (Tobias et al., 2022). Each species was assigned to the diet group that accounted for at least 60 % of its food resources. Species for which no single food type comprised 60 % of the diet were classified as omnivorous. Species groups representing less than 3 % of the community were excluded, yielding four groups for analysis: frugivores, granivores, invertivores, and omnivores.
2.3 Environmental variables
Environmental variables were derived from satellite imagery provided by Bing Maps, using QGIS tools (QGIS.org, 2023), which allowed the consistent and spatially explicit quantification of field- and landscape-scale characteristics across all study sites. These were designed to capture both local (field-scale) and broader (landscape-scale) drivers of bird species richness.
To characterize rice fields, we derived three variables: (1) average rice field width; (2) parcel area, used as a proxy for the effect of bund vegetation; and (3) percentage cover of trees and shrubs. “Field” refers to one entire rice field, which in many cases corresponds to the full extent of the floodplain. “Parcel” refers to a smaller management unit within a field, cultivated independently by one family and delimited by earthen bunds supporting herbaceous and shrub vegetation. Smaller parcels therefore imply a higher perimeter-to-area ratio, resulting in a greater length of bunds per unit area and, consequently, more extensive associated vegetated habitat. Hereafter, this terminology is adopted throughout the text. These variables were selected because they reflect the key structural features of rice fields that can influence bird richness by affecting habitat heterogeneity, edge effects, and resource availability. Field width (m) was measured every 100 m along each transect and averaged. Parcel area (m2) was estimated from a sample of 25 randomly selected parcels in each rice field. Given the low within-field variation in parcel size, any upward bias introduced by the random-point method is expected to be minimal. Tree and shrub cover was estimated by generating 50 random points per rice field and recording whether each fell within woody vegetation. Trees and shrubs were grouped, as satellite imagery did not allow a reliable differentiation. Random points were generated using the “random points in polygons” tool in QGIS.
To characterize the surrounding landscape, we mapped land-use categories (mudflat, open, wooded, forested, and built-up; Table S1 in the Supplement) using manual digitization. Land cover was assessed within buffers in a geometric progression of 100, 200, 400, 800, 1600, and 3200 m around each rice field. These buffers were selected to capture responses across multiple spatial scales while limiting redundancy among highly correlated predictors. They represent spatial scales relevant to the foraging and dispersal capacities of birds (Miguet et al., 2016; Redlich et al., 2018), and to approximate the movement capacity of species with larger home ranges in our dataset (e.g. Ardeidae; Brzorad et al., 2021). The resulting maps were used as input for Fragstats (McGarigal et al., 2023) to calculate (1) landscape diversity (SIDI), which quantifies the compositional heterogeneity of the landscape by accounting for both the richness and relative area of land-cover classes; (2) landscape fragmentation (ENN_MN), which measures the mean Euclidean distance between nearest neighbouring patches of the same class and thus reflects the degree of spatial isolation among habitat patches; and (3) edge complexity (PARA_MN), which captures patch shape complexity by expressing the mean perimeter:area ratio of individual patches, with higher values indicating more irregular and convoluted patch boundaries. These metrics were selected because they represent complementary aspects of landscapes that are ecologically meaningful for patterns of bird richness (Lu et al., 2024).
2.4 Data analysis
To address the objectives of this study, analyses were conducted in three steps: (i) estimation of species richness, (ii) assessment of the effects of individual environmental variables, and (iii) evaluation of their combined influence across spatial scales.
2.4.1 Estimation of species richness
Bird species richness was estimated for each transect survey (transect × date), both overall and for each diet group. Species richness was used as the primary biodiversity metric (q = 0 Hill number), reflecting our focus on patterns of species occurrence and functional representation rather than abundance-weighted community structure. We used a framework robust to small sample sizes (Dorazio et al., 2006; Kéry and Royle, 2008; Yamaura et al., 2016) by fitting Bayesian generalized additive mixed models (GAMMs) in the brms package (Bates et al., 2015; Bürkner, 2017). Because richness is a non-negative count with variance increasing with the mean, observed richness counts were modelled with a Poisson distribution. The total number of individuals was included as a smooth predictor to account for differences in sampling effort among transects, recognizing that it reflects both sampling intensity and underlying variation in abundance, and allowing a flexible (non-linear) relationship with species richness (Chao and Jost, 2012; Gotelli and Colwell, 2001). Month was included as a random effect to account for between-month variation. Although detectability was not explicitly modelled, surveys followed a standardized protocol and were repeated across transects, reducing variability in detection. In addition, the inclusion of the total number of individuals as a proxy for sampling effort partly accounts for differences in detectability among surveys. As our analyses focus on relative patterns across environmental gradients, any remaining detectability bias is expected to have limited influence on the results. Default priors from brms were used, corresponding to weakly informative Student-t priors on intercepts and variance components, and weakly regularizing priors on regression coefficients. These priors provide regularization without constraining parameter estimates and are appropriate for ecological count data and small sample sizes (Bürkner, 2017; Gelman et al., 2008; Tchetgen Tchetgen et al., 2021). This approach estimates expected species richness conditional on sampling effort and detection, rather than using raw observed counts.
For each transect survey, the posterior mean of estimated richness was extracted and used as the response variable in subsequent analyses. This two-stage approach allowed us to separate richness estimation from the evaluation of environmental drivers. Spatial autocorrelation in estimated richness was assessed using Moran's I for both overall and trophic guild richness (Griffith, 2010). No significant autocorrelation was found.
2.4.2 Effects of individual environmental variables
To identify the main drivers of bird richness, we fitted univariate linear mixed models (LMMs) with Gaussian errors, using estimated richness as the response variable and including field-scale (field width, parcel area, and tree cover) and landscape-scale (diversity, fragmentation, and edge complexity) predictors across multiple buffer distances. Month was included as a random effect to account for repeated sampling and to reduce temporal pseudo-replication, thereby leveraging within-month replication to increase the effective sample size without inflating degrees of freedom. Alternative random effects structures (transect, field, and fortnight) were evaluated but did not improve the model fit or affect fixed-effect estimates; we therefore retained month as the sole random effect as a parsimonious representation of temporal dependence. For frugivores, invertivores, and omnivores, richness was log-transformed to reduce the right-skewness and to improve the model fit (Curran-Everett, 2018).
All predictors were standardized (mean centred and scaled by standard deviation). Model assumptions were evaluated with DHARMa residual diagnostics, including the Kolmogorov–Smirnov test for distributional deviations, and a two-sided binomial outlier test (Hartig, 2024).
2.4.3 Integrated effects of field and landscape structure
To assess the combined influence of environmental variables and reduce collinearity, principal component analysis (PCA) was used to summarize correlated field- and landscape-scale descriptors into orthogonal gradients. Variables were rescaled to a 0–1 range (min–max normalization) prior to analysis to avoid differences in measurement scale and enable direct comparison between field- and landscape-scale effects. The first two principal components were retained based on explained variance and interpretability. Variable loadings were used to interpret the contribution of each variable to the resulting gradients. Composite predictors derived from the PCA scores represented field-scale (“field openness”) and landscape-scale (“landscape structure”) gradients.
Univariate LMMs were first fitted for each composite predictor across buffer distances, including month as a random effect to account for temporal dependence. Collinearity between composite predictors was assessed using Spearman's rank correlation. Because field openness and landscape structure were strongly correlated at the 100 m buffer (r = −0.73; Tabachnick et al., 2007), landscape structure at that scale was excluded from the analysis.
For landscape structure, buffer size was selected based on model performance across spatial scales. Specifically, univariate LMMs were fitted for each buffer distance, and the scale included in the multivariate model corresponded to the buffer that minimized the Akaike information criterion corrected for small sample sizes (AICc), thereby identifying the spatial scale at which landscape structure best explained variation in species richness (Burnham and Anderson, 2002).
To assess the relative influence of field- and landscape-scale gradients on species richness, we fitted multivariate LMMs including both composite predictors, again including month as a random effect. Non-linear relationships (quadratic, cubic, and square-root terms) were tested but not retained. Multicollinearity was assessed using variance inflation factors (VIFs). Model assumptions were evaluated using DHARMa diagnostics (Hartig, 2024).
All reported effects and intervals correspond to the linear mixed models (Sect. 2.4.2 and 2.4.3) rather than the Bayesian richness estimation step.
All analyses were conducted in R v. 4.2.1 (R Core Team, 2022a) within RStudio (R Core Team, 2022b). LMMs were fitted with the lme4 package (Brooks et al., 2017). Model selection and validation used MuMIn and car (Bartoń, 2025; Fox and Weisberg, 2018), in addition to DHARMa (Hartig, 2024). Moran's I and PCA were conducted using the spdep and FactoMineR packages, respectively (Bivand and Wong, 2018; Lê et al., 2008). Statistical significance was set at p < 0.05.
A total of 150 bird species, belonging to 53 families and 17 orders, were recorded (Table S2). Estimated species richness per transect survey ranged from 8.5 to 24.8, with a mean of 18.2 species. Among trophic guilds, granivores and invertivores were the most diverse, with averages of 4.9 and 3.7 species, respectively.
3.1 Effects of rice field characteristics on bird communities
The effects of field-scale variables differed between the overall community and trophic guilds (Fig. 2 and Table S3). Variation in parcel area did not affect overall community richness, nor did it influence richness within any trophic guild. In contrast, both tree cover and field width were positively associated with community richness, with increases of approximately 2.9 % per 1 SD. Frugivore richness increased with both tree cover and field width, whereas granivore and omnivore richness responded positively solely to field width, and invertivore richness responded positively solely to tree cover.
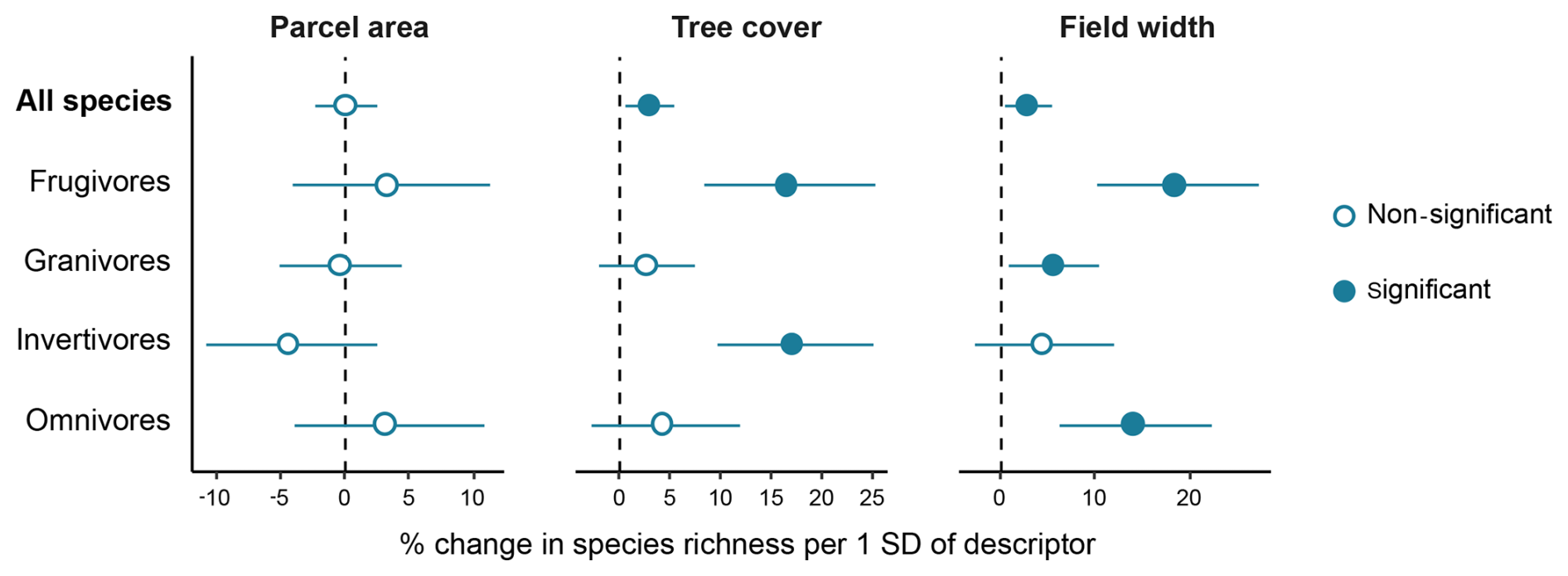
Figure 2Forest plots showing the percentage change in bird species richness per 1 SD increase in descriptors at field scale: parcel area, tree cover, and field width. Circles represent estimated percentage changes and horizontal lines the 95 % intervals.
3.2 Influence of landscape structure on bird communities
Most landscape variables showed clear effects on bird richness, evaluated at the most relevant scale for each group – 800 m for overall and granivore richness, and 1600 m for the remaining groups (Fig. 3 and Table S4). Richness declined with increasing patch fragmentation for all groups, although this effect was uncertain for frugivores and granivores. Landscape diversity was consistently negatively associated with richness, with the strongest declines observed in frugivores (22.6 % per 1 SD) and invertivores (19.2 %). In contrast, edge complexity was positively related to richness, with increases for the community and all trophic guilds.
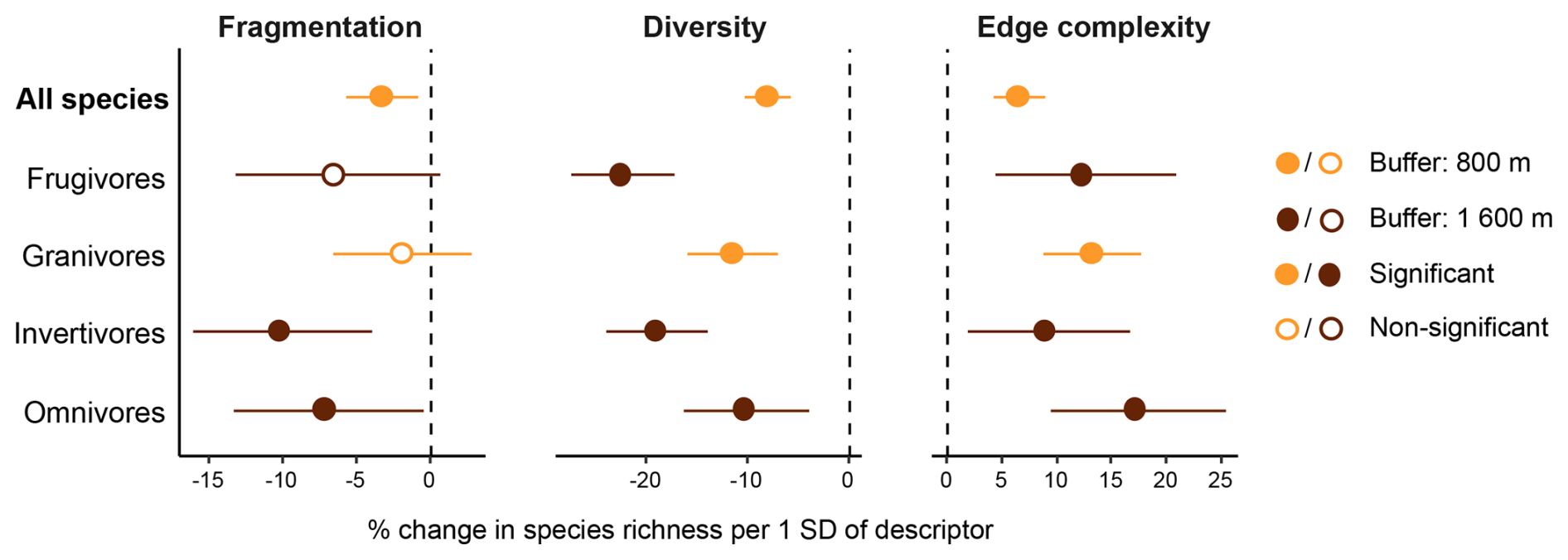
Figure 3Forest plots showing the percentage change in bird species richness per 1 SD increase in landscape metrics: fragmentation, diversity, and edge complexity. Results are shown only for the best buffer models for the overall community and each trophic guild. Circles represent estimated percentage changes and horizontal lines the 95 % intervals.
3.3 Contribution of field- and landscape-scale predictors to bird communities
For the surrounding landscape, we performed PCAs at all buffer distances (200–3200 m; Figs. 4b, c and S2). Model selection based on AICc identified 800 m as the most relevant scale for overall and granivore richness, and 1600 m for frugivore, invertivore, and omnivore richness, with these models showing the lowest AICc values and consistent support relative to alternative buffer sizes (Table S5).
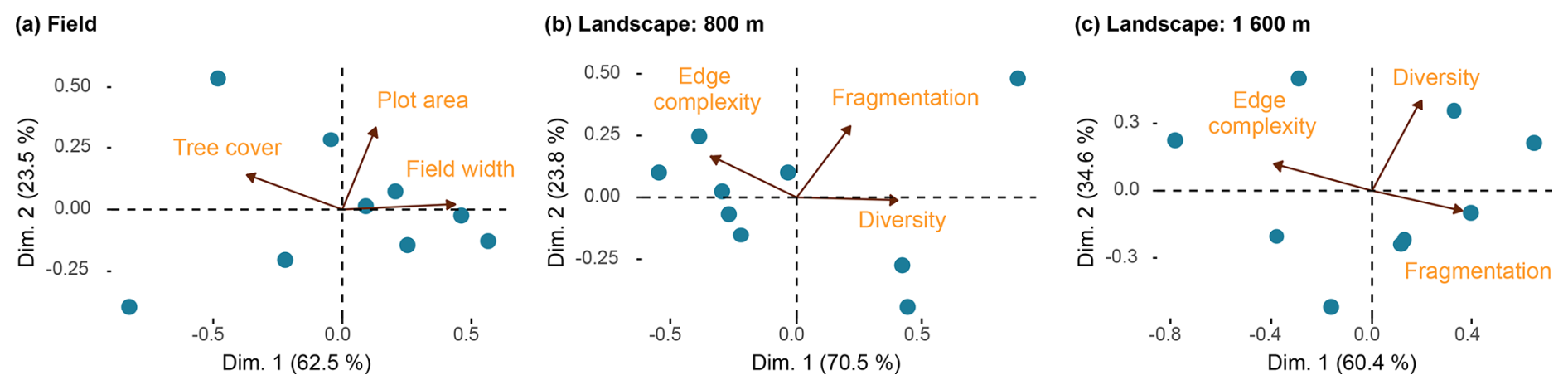
Figure 4Principal component analysis (PCA) summarizing field (a) and landscape variables measured at 800 m (b) and 1600 m (c). The proportion of variance explained by the first two components (Dim. 1 and Dim. 2) is indicated in parentheses. Arrows show the direction and relative contribution of each variable, and blue dots represent sampled sites.
The PCA of field characteristics summarized correlated variables into two main gradients (Fig. 4a and Table S6). The first principal component (62.5 % variance) reflects a gradient contrasting field width and tree cover, while the second component (23.5 %) is associated with parcel area. The resulting composite variable, “field openness”, captures a gradient from narrow, tree-rich fields (negative values) to wide, open fields with sparse trees (positive values), and was used to assess the combined influence of field-scale structure (see Table S7 for LMM results of “field openness”).
Similarly, PCAs of landscape variables at the selected spatial scales summarized multivariate structure into dominant gradients (Fig. 4b, c and Table S6). At both 800 and 1600 m, the first principal component described a gradient opposing landscapes with more complex patch shapes to those that are more diverse and fragmented. The composite variable “landscape structure” therefore represents structurally complex, less fragmented landscapes (negative values) to more heterogeneous but fragmented landscapes (positive values). This composite predictor was used to assess the integrated effects of landscape structure across scales.
Species richness per transect survey declined with “landscape structure” (Fig. 5a and Table S8) for all groups. “Field openness” did not influence overall or granivore richness, but it had a positive effect on frugivore, invertivore, and omnivore richness (Fig. 5b–e and Table S8). The apparent low species numbers estimated in the GAMMs reflect that each value corresponds to the richness recorded in a single visit to a transect, rather than a transect-level average. Overall, the effect sizes associated with landscape structure were consistently larger and more significant than those of field openness. This contrast is evident in the steeper slopes and higher significance of landscape structure compared to field openness across models. Because all predictors were standardized, these differences are directly comparable, indicating a stronger influence of landscape-scale factors on species richness (Fig. 5 and Table S8).
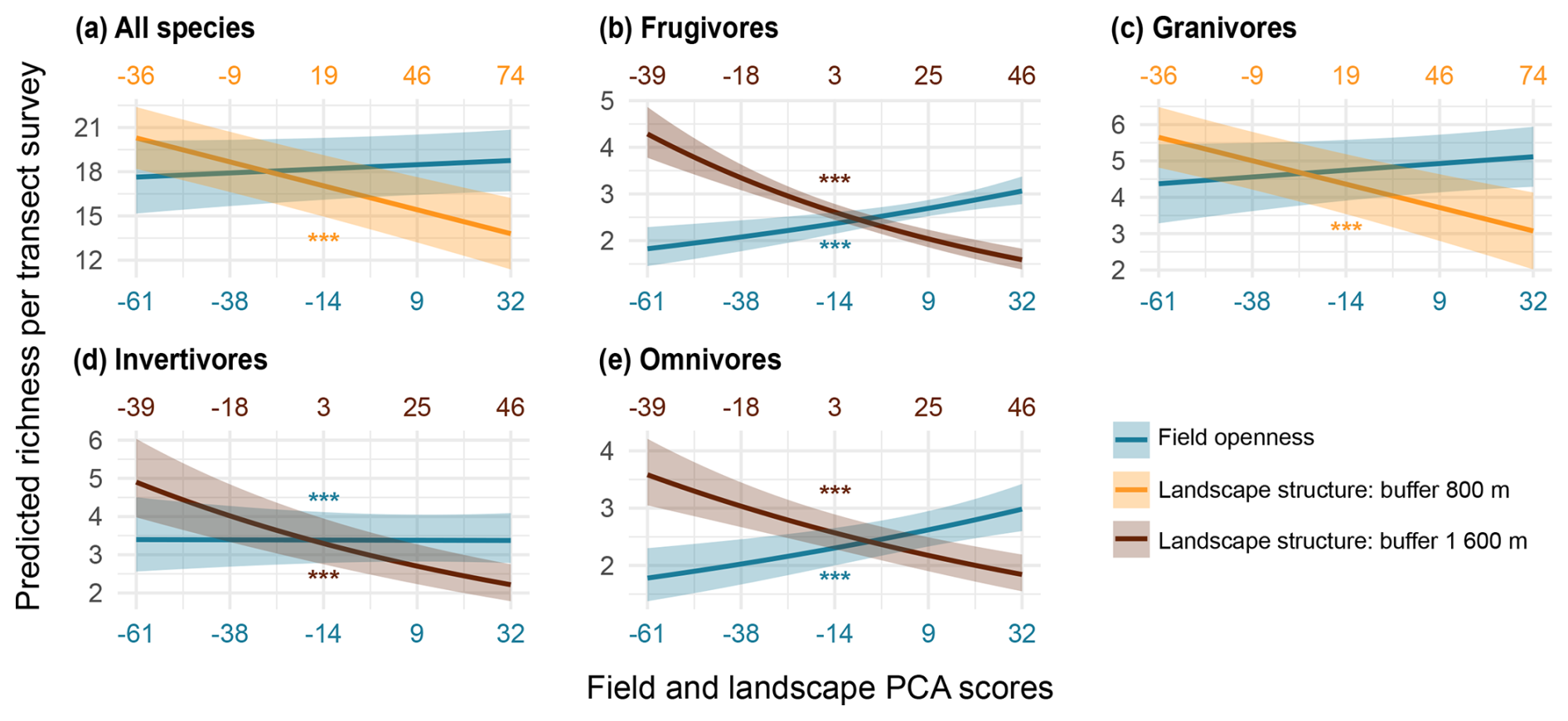
Figure 5Linear mixed models (LMMs) showing predicted bird species richness per transect survey as a function of field openness and landscape structure for all species (a) and trophic guilds (b–e). Shaded areas represent 95 % confidence intervals. Month was included as a random effect. Significance level: = p < 0.001. Field PCA scores range from narrow, tree-rich fields (negative values) to wide, open fields with sparse tree cover (positive values). Landscape PCA scores range from landscapes with intricate, well-connected patch shapes (negative values) to those that are more compositionally diverse but also more fragmented (positive values).
Our study provides one of the first evaluations of the drivers of bird species richness in smallholder African rice agroecosystems, a system that remains central to local food production yet is poorly represented in ecological research. Bird richness responded to both field- and landscape-scale characteristics, with landscape-scale gradients showing consistently stronger associations across trophic guilds. Field-level effects were more context dependent, with “field openness” having limited influence overall but a positive effect for some guilds. At the landscape scale, bird richness declined along the gradient of increasing “landscape structure”, indicating that more fragmented and compositionally diverse landscapes were associated with lower richness. These gradients reflect underlying relationships with individual environmental descriptors. For example, the positive responses of some guilds to field openness are consistent with associations with field width and tree cover, while the negative relationship with landscape structure reflects the combined effects of fragmentation and landscape diversity. Trophic guilds also differed in their scale of response, consistent with differences among functional groups in mediating sensitivity to habitat structure (Jackson and Fahrig, 2015; Miguet et al., 2016).
4.1 Effects of rice field characteristics on bird communities
Field-scale vegetation structure emerged as an important driver of bird community richness, particularly across trophic guilds. Scattered trees and shrubs increased structural heterogeneity in fields and may facilitate the use of fields by species that rely on woody elements across the landscape (Lee et al., 2024; Martin et al., 2009). Rice fields containing woody vegetation supported higher invertivore richness, as trees provide additional foraging substrates and microhabitats, a pattern also observed for insectivorous bats in this region (Fernandes et al., 2025). Homogeneous fields without trees can host some foliage-gleaning invertivores adapted to open herbaceous vegetation (Bos et al., 2006). However, the presence of scattered trees broadens the available foraging niches by attracting bark probers, hover-gleaners, and perch-hunting species (Hološková et al., 2025). One widely occurring species, the winter thorn (Faidherbia albida), is recognized locally and in the scientific literature for supporting diverse arthropod communities (Zwarts et al., 2023). Frugivorous birds similarly benefited from the presence of trees, likely reflecting the availability of fruit resources. Fruit-producing species such as the giant-leaved fig tree (Ficus lutea), an important component of West African trophic networks (Daru et al., 2015), occur in the study area, although its specific contribution in our system remains untested.
Trees and shrubs thus performed multiple ecological functions in fields by providing refuge, nesting sites, perching structures, and a wider range of food resources (Angkaew et al., 2023; Hološková et al., 2025). However, contrary to our expectations, parcel area, used here as a proxy for the extent of natural vegetation cover along bunds, did not influence bird richness. This suggests that not all aspects of field-scale structural heterogeneity contribute equally to bird richness and that the ecological relevance of such features depends on their magnitude and functional expression (Fahrig, 2017).
Field width also influenced bird species richness. Within the heterogeneous smallholder landscape, wider rice fields supported greater overall bird richness. This pattern likely reflects moderate increases in field area within a structurally diverse mosaic, rather than the homogenizing effect typically associated with large monocultures, which are absent in this system (Zhu et al., 2024). Because fields in this system are relatively narrow, increases in width expand the effective foraging area without eroding fine-scale structural diversity, which is characteristic of traditional rice landscapes (Benton et al., 2021; He et al., 2023; Zhu et al., 2024). Granivores and omnivores, which are typically opportunistic groups (Buelow and Sheaves, 2015; Howe and Brown, 1999), responded positively to field width, consistent with their preference for open habitats where grains and other accessible resources accumulate (Katuwal et al., 2022). Similar responses have been reported in Asian rice systems (e.g. Amano et al., 2008; Son et al., 2024), and our results show that this field-scale mechanism also applies to smallholder floodplain rice fields in West Africa. Frugivores showed the strongest positive response to field width, with wider rice fields, which are typically more open, likely increasing the visibility and accessibility of fruiting trees within or adjacent to fields (Tscharntke et al., 2008).
Taken together, these results reveal a complementary pattern at the field scale: openness favoured granivorous and omnivorous species, which tend to exploit predictable resources in open fields (Buelow and Sheaves, 2015; Voudouri et al., 2021). In contrast, scattered trees supported frugivores and invertivores whose diets depend on more complex and temporally variable trophic interactions, such as fruiting events or arthropod dynamics (Daru et al., 2015; França et al., 2020). This balance between open foraging areas and woody elements likely contributes to maintaining functional diversity within smallholder rice systems.
4.2 Influence of landscape structure on bird communities
Landscape characteristics strongly influenced bird species richness, with clear differences among trophic guilds and spatial scales. The scale of response reflected how mobility and resource distribution shape species' sensitivity to habitat structure (Rolando, 2002). Frugivore richness was primarily driven by landscape composition at broader spatial scales (1600 m around rice fields), consistent with the dispersed distribution of fruiting resources in tropical landscapes (Blendinger et al., 2015). Invertivores also showed broad-scale responses, possibly because they are forced to range widely to locate prey with naturally dynamic and not reliable distributions (França et al., 2020). Omnivores, which rely on a wide variety of patchily distributed resources, also responded to landscape attributes at broader scales, in line with their flexible habitat use in heterogeneous mosaics (Habel et al., 2019).
More fragmented landscapes supported fewer bird species, suggesting that increased fragmentation may constrain movement and access to complementary habitats. Invertivores, in particular, are known to be sensitive to habitat fragmentation and often rely on movement across connected areas to track ephemeral prey (Otieno and Mukasi, 2023). Empirical evidence further shows that smaller or more isolated habitat patches tend to support individuals in poorer conditions or younger cohorts, suggesting that high-quality territories are concentrated in more continuous habitat (Coddington et al., 2023; Luther et al., 2024). Although connectivity generally promotes species richness by facilitating movement and access to complementary resources (Cushman et al., 2013), some groups, such as granivores and frugivores, may be less dependent on landscape corridors when local resources (e.g. spilled grains, fruiting trees) are abundant (Estrada et al., 1993; van Osta et al., 2024). This suggests that, in relatively permeable smallholder landscapes, local resource availability can outweigh spatial continuity for certain trophic guilds.
Landscape diversity had a negative effect on bird richness. Although high landscape diversity is often considered beneficial because it reflects habitat variety (Matuoka et al., 2020), in this region, higher diversity values may be associated with mosaics shaped by human land-use practices, such as settlement expansion, selective tree removal, and shifting cultivation. In such contexts, increases in compositional heterogeneity can coincide with structural fragmentation and reduced habitat continuity, potentially contributing to lower bird richness.
Edge habitats positively influenced bird richness in rice fields at both the community and dietary group levels. Ecotones offer abundant foraging opportunities and increase structural diversity, attracting species from multiple guilds (Berry, 2001). Rice fields adjacent to complex landscape patches likely possess more irregular boundaries, increasing habitat interface length and facilitating the spillover of birds and resources across habitats (Berry, 2001). However, the ecological benefits of edges in bird richness are context dependent, and positive responses likely arise from edges adjoining semi-natural vegetation rather than from edges bordering highly modified or built-up areas (Karimnejad et al., 2024).
Our results extend previous evidence on the role of landscape configuration in shaping bird richness in agricultural mosaics (e.g. Amano et al., 2008; Cabral et al., 2021; Hološková et al., 2025; Priyadarshana et al., 2024; Stafford et al., 2010). They reveal that trophic guilds differ markedly in their scale of response and in the landscape attributes most relevant to them, highlighting how differences in functional groups and mobility constraints mediate biodiversity patterns in smallholder West African rice farming landscapes. This underscores the need for managing the landscape matrix to support bird diversity. In traditional smallholder systems, sustaining bird communities depends on preserving diverse yet connected habitat networks rather than relying exclusively on field-level management interventions in isolation from the broader landscape context.
4.3 Relative importance of field and landscape traits
When field- and landscape-scale characteristics were considered together, bird richness showed stronger associations with landscape-scale gradients than with field-scale characteristics, both overall and within trophic guilds. This pattern suggests that, in smallholder low-intensity systems, local management operates within constraints imposed by the broader landscape configuration, which can influence species availability and movement.
Bird richness declined in landscapes characterized by greater habitat diversity, higher fragmentation, and more regular patch shapes. Although these results contrast with findings from other regions (e.g. Hovick et al., 2015; Lee et al., 2024; Leyequién et al., 2010), they are consistent with landscape patterns in the study region, where higher landscape diversity values may reflect human-modified mosaics. In such contexts, increases in compositional diversity can coincide with fragmentation and reduced habitat continuity, potentially contributing to lower bird richness. Moreover, landscape heterogeneity is not universally beneficial (King et al., 2010), as some open-country species depend on extensive, continuous habitats. Additionally, proximity to certain patches, such as forest remnants, may impose higher predation risk due to increased densities of avian and mammalian predators (Renfrew et al., 2005).
Field openness did not affect overall or granivore richness. Although trees are important for frugivores, frugivore richness was higher in more open fields. Part of this pattern may reflect a slight detectability bias, as all bird groups are generally easier to observe in open habitats (Sólymos et al., 2018), but frugivores may also take advantage of the greater visibility and accessibility of fruiting trees located within or adjacent to more open fields (Tscharntke et al., 2008). Invertivores showed a small positive response to field openness, suggesting that they can exploit resources in open areas despite their reliance on trees and shrubs (Castaño-Villa et al., 2014; Khamcha et al., 2018). Rice fields often contain high densities of exposed invertebrates after ploughing, providing short-term foraging opportunities that may benefit invertivores (Katuwal et al., 2022).
The stronger influence of landscape-scale features over field-level characteristics on bird species richness aligns with evidence from other tropical agricultural systems, where landscape context, particularly the amount and configuration of surrounding vegetation, is a key determinant of avian diversity in farmland habitats (Jarrett et al., 2021). Birds are highly mobile organisms that exploit resources across broad spatial extents, and local habitats such as individual rice fields likely represent only a fraction of the area used by most species; the surrounding landscape thus shapes the regional species pool from which local communities are assembled.
The positive response of granivores to landscape structure was unexpected, given that this guild is generally considered more tolerant of habitat disturbance and less dependent on landscape complexity than frugivores or invertivores (Bregman et al., 2016; Jarrett et al., 2021). However, the rice agroecosystem studied here reflects a long-established, heterogeneous agricultural mosaic rather than a forest-to-agriculture conversion gradient. In this context, granivores may rely on landscape mosaics to access complementary seed resources, rendering their richness sensitive to landscape features. Notably, granivores responded at a finer spatial scale (800 m) than frugivores and invertivores (1600 m), suggesting that while all guilds are influenced by landscape structure, granivores integrate resources over shorter distances, consistent with their generally lower habitat specialization.
4.4 Limitations and future work
Some methodological constraints should be considered when interpreting our findings. Bird surveys conducted in open environments such as wider rice fields may be subject to detection biases, as visually conspicuous species may be recorded more readily than canopy-associated ones in tree-covered plots. Nevertheless, the use of Bayesian species richness estimators, which explicitly account for imperfect detection, likely reduced this bias. The study was based on nine transects, which limits spatial replication and statistical power. However, repeated sampling across two rice-growing cycles, combined with the inclusion of a temporal random effect, provided sufficient observations to detect consistent associations between bird richness and environmental predictors. Environmental variables were derived from remotely sensed imagery, which provides a consistent and spatially explicit characterization of field- and landscape-scale structure but may overlook fine-scale habitat features (e.g. vegetation structure along bunds) that could influence bird richness. Incorporating ground-based habitat measurements would help to refine these relationships in future studies.
The assignment of species to trophic guilds represents a simplification. Many birds exhibit flexible feeding behaviours, and their classification into discrete guilds may obscure finer ecological responses to habitat gradients.
Future studies should explore additional field characteristics, such as field area, water depth, permanence, and the extent of uncultivated parcels, to clarify their combined influence on bird species richness. Examining these variables jointly would be particularly informative, although it would require increasing the number of surveyed rice fields to ensure sufficient statistical power. Further work could also test whether the relative influence of landscape and field characteristics varies across years or under climatic extremes. In unusually dry or wet years, field-scale characteristics may become more influential, as local microhabitat conditions may be critical for bird persistence (King et al., 2010). Long-term monitoring would help to clarify how inter-annual climatic variation mediates the balance between local and landscape drivers of bird species richness. Because species richness gives equal weight to all species, incorporating abundance-weighted diversity metrics (e.g. Hill numbers with q > 0) would provide complementary insights into the community structure and dominance patterns.
4.5 Conclusions and management implications
Our results show that bird species richness in smallholder rice fields is more strongly associated with the configuration of the surrounding landscape than with field-scale characteristics, highlighting the importance of considering rice fields as components of broader agricultural mosaics. In this context, landscape configuration influences how birds access resources distributed across space and shapes the communities that can persist locally. Differences among trophic guilds further indicate that functional characteristics mediate sensitivity to spatial scale and habitat structure, reinforcing the value of functional approaches for understanding biodiversity patterns in agroecosystems.
Although agri-environmental schemes and governmental policies remain limited in developing countries due to financial constraints (Wright et al., 2012), our findings suggest that integrative land-use planning can play an important role in reconciling food production with biodiversity conservation. In agricultural mosaics, conservation strategies are likely to benefit from landscape-level approaches that limit excessive habitat fragmentation while maintaining edge complexity, thereby supporting a connected matrix capable of sustaining diverse bird communities (Berry, 2001; He et al., 2023). Local management actions can complement these broader measures. Retaining and planting scattered trees and shrubs within rice fields enhances habitat quality for invertivores, which contribute pest control services that may reduce reliance on chemical inputs (Díaz-Siefer et al., 2022).
More broadly, this study highlights the need to interpret commonly used heterogeneity metrics in light of their spatial configuration and ecological context, rather than assuming uniformly positive effects. By linking multi-scale drivers of bird richness with diet-based functional groupings, our results contribute to a more nuanced understanding of how landscape structure shapes biodiversity in smallholder agricultural systems, with implications for both ecological research and agroecological management in tropical regions.
The data and code necessary to reproduce the results of this paper are publicly available on the Open Science Framework (OSF) at https://doi.org/10.17605/OSF.IO/TRFNQ (Lopes et al., 2026). This repository includes estimated species richness per transect survey, transect-level field variables, landscape metrics, and the full R script used for all analyses. Raw transect survey data are part of an ongoing PhD project and will be made publicly available upon its completion; in the meantime, they can be shared upon reasonable request.
The supplement related to this article is available online at https://doi.org/10.5194/we-26-103-2026-supplement.
PL – methodology, investigation, data curation, formal analysis, visualization, and writing (original draft, review, and editing). IL – methodology, investigation, data curation, and writing (review and editing). PC – methodology, investigation, and writing (review and editing). RL – investigation and writing (review and editing). MC – investigation and writing (review and editing). ML – methodology, investigation, validation, and writing (review and editing). JMP – funding acquisition, supervision, validation, and writing (review and editing). AR – conceptualization, funding acquisition, project administration, methodology, supervision, validation, and writing (review and editing).
The contact author has declared that none of the authors has any competing interests.
Publisher's note: Copernicus Publications remains neutral with regard to jurisdictional claims made in the text, published maps, institutional affiliations, or any other geographical representation in this paper. The authors bear the ultimate responsibility for providing appropriate place names. Views expressed in the text are those of the authors and do not necessarily reflect the views of the publisher.
We would like to foremost thank the local communities and farmers who welcomed us into their fields and allowed for this research to take place. During the field work, we had the collaboration and logistic support of the KAFO Peasant Federation in Guinea-Bissau. We are especially grateful to its secretary-general, Sambú Seck, for his support. We thank all the staff of KAFO's Djalicunda Center, particularly those who facilitated our day-to-day activities and provided support at all levels. Finally, we thank the reviewers for their careful reading of the paper and for their constructive and insightful comments, which substantially improved the quality of this work.
This study was funded by Fundação para a Ciência e Tecnologia, I.P./MCTES through national funds (PIDDAC) via project PTDC/ASPAGR/0876/2020 (https://doi.org/10.54499/PTDC/ASP-AGR/0876/2020), PhD grant to PL (022.14253.BD; https://doi.org/10.54499/2022.14253.BD), PhD grant to PC (PD/BD/150566/2020; https://doi.org/10.54499/PD/BD/150566/2020), and structural funds to CE3C (UID/BIA/00329/2023). Prepared in part while AR was funded by ERC-ABIDE (CoG ID 101043231). PL was also funded by a grant from FCiências.ID – Associação para a Investigação e Desenvolvimento de Ciência.
This paper was edited by Ricardo Rocha and reviewed by Luke Powell, Ariane Dellavalle, and two anonymous referees.
Amano, T. and Yamaura, Y.: Ecological and life-history traits related to range contractions among breeding birds in Japan, Biol. Conserv., 137, 271–282, https://doi.org/10.1016/j.biocon.2007.02.010, 2007.
Amano, T., Kusumoto, Y., Tokuoka, Y., Yamada, S., Kim, E. Y., and Yamamoto, S.: Spatial and temporal variations in the use of rice-paddy dominated landscapes by birds in Japan, Biol. Conserv., 141, 1704–1716, https://doi.org/10.1016/j.biocon.2008.04.012, 2008.
Angkaew, R., Ngoprasert, D., Powell, L. A., Limparungpatthanakij, W., Round, P. D., and Gale, G. A.: Variation in avian responses to rice intensification in the central plains of Thailand guides conservation opportunities, Biol. Conserv., 286, 110307, https://doi.org/10.1016/j.biocon.2023.110307, 2023.
Bartoń, K.: MuMIn: Multi-Model Inference, https://cran.r-project.org/package=MuMIn (last access: 19 May 2025), 2025.
Bates, D., Martin, M., Bolker, B., and Walker, S.: Fitting Linear Mixed-Effects Models Using lme4, J. Stat. Softw., 67, 1–48, https://doi.org/10.18637/jss.v067.i01, 2015.
Benton, T., Bieg, C., Harwatt, H., Pudassaini, R., and Wellesley, L.: Food system impacts on biodiversity loss: Three levers for food system transformation in support of nature, Research report, Chatham House, London, 1–71, ISBN 978 1 78413 433 4, 2021.
Berry, L.: Edge effects on the distribution and abundance of birds in a southern Victorian forest, Wildl. Res., 28, 239–245, https://doi.org/10.1071/WR00057, 2001.
Bibby, C., Burgess, N., Hill, D., and Mustoe, S.: Bird census techniques, Elsevier, London, ISBN 978-0-12-095831-3, 2000.
Bivand, R. S. and Wong, D. W. S.: Comparing implementations of global and local indicators of spatial association, Test, 27, 716–748, https://doi.org/10.1007/s11749-018-0599-x, 2018.
Blendinger, P. G., Jiménez, J., Macchi, L., Martín, E., Sánchez, M. S., and Ayup, M. M.: Scale-dependent spatial match between fruits and fruit-eating birds during the breeding season in Yungas Andean forests, Biotropica, 47, 702–711, https://doi.org/10.1111/btp.12247, 2015.
Bos, D., Grigoras, I., and Ndiaye, A.: Land cover and avian biodiversity in rice fields and mangroves of West Africa, Wetlands International, Dakar, ISBN 978-9-05-882034-1, 2006.
Bregman, T. P., Lees, A. C., MacGregor, H. E. A., Darski, B., de Moura, N. G., Aleixo, A., Barlow, J., and Tobias, J. A.: Using avian functional traits to assess the impact of land-cover change on ecosystem processes linked to resilience in tropical forests, P. R. Soc. B, 283, 20161289, https://doi.org/10.1098/rspb.2016.1289, 2016.
Brooks, M. E., Kristensen, K., van Benthem, K. J., Magnusson, A., Berg, C. W., Nielsen, A., Skaug, H. J., Mächler, M., and Bolker, B. M.: glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling, R J., 9, 378–400, https://doi.org/10.32614/rj-2017-066, 2017.
Brzorad, J. N., Allen, M. C., Jennings, S., Condeso, E., Elbin, S., Kays, R., Lumpkin, D., Schweitzer, S., Tsipoura, N., and Maccarone, A. D.: Seasonal Patterns in Daily Flight Distance and Space Use by Great Egrets (Ardea alba), Waterbirds, 44, 343–355, https://doi.org/10.1675/063.044.0309, 2021.
Buelow, C. and Sheaves, M.: A birds-eye view of biological connectivity in mangrove systems, Estuar. Coast. Shelf S., 152, 33–43, https://doi.org/10.1016/j.ecss.2014.10.014, 2015.
Bürkner, P.-C.: brms: An R Package for Bayesian Multilevel Models Using Stan, J. Stat. Softw., 80, 1–28, https://doi.org/10.18637/jss.v080.i01, 2017.
Burnham, K. P. and Anderson, D. R.: Model selection and multimodel inference: a practical information-theoretic approach, 2nd Edn., Springer, New York, ISBN 978-0-387-95364-9, 2002.
Burns, F., Eaton, M. A., Burfield, I. J., Klvaňová, A., Šilarová, E., Staneva, A., and Gregory, R. D.: Abundance decline in the avifauna of the European Union reveals cross-continental similarities in biodiversity change, Ecol. Evol., 11, 16647–16660, https://doi.org/10.1002/ece3.8282, 2021.
Cabral, J. P., Faria, D., and Morante-Filho, J. C.: Landscape composition is more important than local vegetation structure for understory birds in cocoa agroforestry systems, Forest Ecol. Manag., 481–118704, https://doi.org/10.1016/j.foreco.2020.118704, 2021.
Chao, A. and Jost, L.: Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size, Ecology, 93, 2533–2547, https://doi.org/10.1890/11-1952.1, 2012.
Cao, L. and Fox, A. D.: Birds and people both depend on China's wetlands, Nature, 460, 173, https://doi.org/10.1038/460173b, 2009.
Castaño-Villa, G. J., Ramos-Valencia, S. A., and Fontúrbel, F. E.: Fine-scale habitat structure complexity determines insectivorous bird diversity in a tropical forest, Acta Oecol., 61, 19–23, https://doi.org/10.1016/j.actao.2014.10.002, 2014.
Catarino, L., Martins, E. S., Pinto Basto, M. F., and Diniz, M. A.: An annotated checklist of the vascular flora of Guinea-Bissau (West Africa), Blumea: J. Plant Taxon. Plant Geogr., 53, 1–222, https://doi.org/10.3767/000651908X608179, 2008.
Chaves, P., Schaafsma, M., Dabo, D., Lomba, J. Z., Mane, F., de Lima, R. F., Palmeirim, J. M., Rocha, R., Seck, S., Biai, J., Timóteo, S., Meyer, C. F. J., and Rainho, A.: Friend or foe? Attitudes of rice farmers toward wild animals in West Africa, Ecol. Soc., 29, 24, https://doi.org/10.5751/ES-15486-290424, 2024.
Coddington, C. P. J., Cooper, W. J., and Luther, D. A.: Effects of forest fragmentation on avian breeding activity, Conserv. Biol., 37, e14063, https://doi.org/10.1111/cobi.14063, 2023.
Curran-Everett, D.: Explorations in statistics: The log transformation, Adv. Physiol. Educ., 42, 343–347, https://doi.org/10.1152/ADVAN.00018.2018, 2018.
Cushman, S. A., McRae, B., Adriaensen, F., Beier, P., Shirley, M., and Zeller, K.: Biological corridors and connectivity, in: Key Topics in Conservation Biology, vol. 2, edited by: Macdonald, D. W. and Willis, K. J., John Wiley & Sons, Ltd, ISBN 978-1-40-512249-8, 2013.
Daru, B., Yessoufou, K., Nuttman, C., and Abalaka, J.: A preliminary study of bird use of fig Ficus species in Amurum Forest Reserve, Nigeria, Malimbus, 37, 1–15, 2015.
Díaz-Siefer, P., Olmos-Moya, N., Fontúrbel, F. E., Lavandero, B., Pozo, R. A., and Celis-Diez, J. L.: Bird-mediated effects of pest control services on crop productivity: a global synthesis, J. Pest Sci., 95, 567–576, https://doi.org/10.1007/s10340-021-01438-4, 2022.
Dorazio, R. M., Royle, J. A., Söderström, B., and Glimskär, A.: Estimating species richness and accumulation by modeling species occurrence and detectability, Ecology, 87, 842–854, https://doi.org/10.1890/0012-9658(2006)87[842:ESRAAB]2.0.CO;2, 2006.
dos Reis-Silva, F., Pizzigalli, C., Seck, S., Cabeza, M., Rainho, A., Rocha, R., and Palmeirim, A. F.: Unveiling how herpetofauna cope with land-use changes – Insights from forest-cashew-rice landscapes in West Africa, Biotropica, 57, e13416, https://doi.org/10.1111/btp.13416, 2025.
Elphick, C. S.: Landscape effects on waterbird densities in California rice fields: Taxonomic differences, scale-dependence, and conservation implications, Waterbirds, 31, 62–69, https://doi.org/10.1675/1524-4695(2008)31[62:LEOWDI]2.0.CO;2, 2008.
Estrada, A., Coates-Estrada, R., Meritt, D., Montiel, S., and Curiel, D.: Patterns of frugivore species richness and abundance in forest islands and in agricultural habitats at Los Tuxtlas, Mexico, Vegetatio, 107, 245–257, https://doi.org/10.1007/BF00052227, 1993.
Fahrig, L.: Ecological responses to habitat fragmentation per se, Annu. Rev. Ecol. Evol. S., 48, 1–23, https://doi.org/10.1146/annurev-ecolsys-110316-022612, 2017.
Fahrig, L., Baudry, J., Brotons, L., Burel, F. G., Crist, T. O., Fuller, R. J., Sirami, C., Siriwardena, G. M., and Martin, J.-L.: Functional landscape heterogeneity and animal biodiversity in agricultural landscapes, Ecol. Lett., 14, 101–112, https://doi.org/10.1111/j.1461-0248.2010.01559.x, 2011.
FAO: Land statistics 2001-2022: Global, regional and country trends, FAOSTAT Analytical Briefs, No. 88, FAO, https://doi.org/10.4060/cd1484en, 2024.
Fasola, M. and Ruiz, X.: The value of rice fields as substitutes for natural wetlands for waterbirds in the Mediterranean Region, Waterbirds, 19, 122–128, https://doi.org/10.2307/1521955, 1996.
Fernandes, G., Massaad, M., Chaves, P. A., and Rainho, A.: Tree and landscape characteristics outweigh insect abundance in driving bat activity in West African rice fields, Agr. Ecosyst. Environ., 193, 109774, https://doi.org/10.1016/j.agee.2025.109774, 2025.
Fox, J. and Weisberg, S.: Visualizing Fit and Lack of Fit in Complex Regression Models with Predictor Effect Plots and Partial Residuals, J. Stat. Softw., 87, 1–27, https://doi.org/10.18637/jss.v087.i09, 2018.
França, L. F., Figueiredo-Paixão, V. H., Duarte-Silva, T. A., and Dos Santos, K. B.: The effects of rainfall and arthropod abundance on breeding season of insectivorous birds, in a semi-arid neotropical environment, Zoologia, 37, 1–7, https://doi.org/10.3897/zoologia.37.e37716, 2020.
Garrity, D. P., Akinnifesi, F. K., Ajayi, O. C., Weldesemayat, S. G., Mowo, J. G., Kalinganire, A., Larwanou, M., and Bayala, J.: Evergreen agriculture: a robust approach to sustainable food security in Africa, Food Secur., 2, 197–214, https://doi.org/10.1007/S12571-010-0070-7, 2010.
Gelman, A., Jakulin, A., Pittau, M. G., and Su, Y.-S.: A weakly informative default prior distribution for logistic and other regression models, Ann. Appl. Stat., 2, 1360–1383, https://doi.org/10.1214/08-AOAS191, 2008.
Global Rice Science Partnership: Rice almanac, 4th Edn., International Rice Research Institute, Los Baños, Philipines, ISBN 978-9-71-220300-8, 2013.
Gotelli, N. J., and Colwell, R. K.: Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness, Ecol. Lett., 4, 379–391, https://doi.org/10.1046/j.1461-0248.2001.00230.x, 2001.
Griffith, D. A.: The Moran coefficient for non-normal data, J. Stat. Plan. Inference, 140, 2980–2990, https://doi.org/10.1016/j.jspi.2010.03.045, 2010.
Habel, J. C., Tobias, J. A., and Fischer, C.: Movement ecology of Afrotropical birds: Functional traits provide complementary insights to species identity, Biotropica, 51, 894–902, https://doi.org/10.1111/btp.12702, 2019.
Hartig, F.: DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models, CRAN (The Comprehensive R Archive Network), https://doi.org/10.32614/CRAN.package.DHARMa, 2024.
He, K., Song, A., Zhang, Z., Ramdat, N., Wang, J., Wu, W., and Chen, X.: Restored coastal wetlands with low degree of separation and high patch connectivity attract more birds, Front. Mar. Sci., 10, 1081827, https://doi.org/10.3389/fmars.2023.1081827, 2023.
Hološková, A., Hanzelka, J., Grünwald, J., Szarvas, F., Řeřicha, M., Štrobl, M., Kadlec, T., and Reif, J.: Predictors of farmland bird species richness in intensively used agricultural landscapes: habitat heterogeneity, crop type, and food supply matter, Landsc. Ecol., 40, e03327, https://doi.org/10.1007/s10980-025-02079-z, 2025.
Hovick, T. J., Elmore, R. D., Fuhlendorf, S. D., Engle, D. M., and Hamilton, R. G.: Spatial heterogeneity increases diversity and stability in grassland bird communities, Ecol. Appl., 25, 662–672, https://doi.org/10.1890/14-1067.1, 2015.
Howe, H. F. and Brown, J. S.: Effects of birds and rodents on synthetic tallgrass communities, Ecology, 80, 1776–1781, https://doi.org/10.1890/0012-9658(1999)080[1776:EOBARO]2.0.CO;2, 1999.
Ishong, J. A., Afrifa, J. K., Iwajomo, S. B., Deikumah, J. P., Ivande, S. T., and Cresswell, W.: Population trends of resident and migrant West African bird species monitored over an 18-year period in central Nigeria, Ostrich, 93, 171–186, https://doi.org/10.2989/00306525.2022.2068691, 2022.
Jackson, H. B. and Fahrig, L.: Are ecologists conducting research at the optimal scale?, Global Ecol. Biogeogr., 24, 52–63, https://doi.org/10.1111/geb.12233, 2015.
Jarrett, C., Smith, T. B., Claire, T. T. R., Ferreira, D. F., Tchoumbou, M., Elikwo, M. N. F., Wolfe, J., Brzeski, K., Welch, A. J., Hanna, R., and Powell, L. L.: Bird communities in African cocoa agroforestry are diverse but lack specialized insectivores, J. Appl. Ecol., 58, 1237–1247, https://doi.org/10.1111/1365-2664.13864, 2021.
Jiang, J., Zhang, H., Ge, J., Zuo, L., Xu, L., Song, M., Ding, Y., Xie, Y., and Huang, W.: The 20 m Africa rice distribution map of 2023, Earth Syst. Sci. Data, 17, 1781–1805, https://doi.org/10.5194/essd-17-1781-2025, 2025.
Karimnejad, M., Malekian, M., Pourmanafi, S., Mobarakeh, Z. M., Keramati, S., Ghased, R., and Ahmadi, M.: Forest edge encroachment by rural orchards shifts bird communities in favor of understory birds: Forest birds as indicators of landscape changes in agroecosystems, Ecol. Indic., 167, 112698, https://doi.org/10.1016/j.ecolind.2024.112698, 2024.
Katuwal, H. B., Rai, J., Tomlinson, K., Rimal, B., Sharma, H. P., Baral, H. S., Hughes, A. C., and Quan, R. C.: Seasonal variation and crop diversity shape the composition of bird communities in agricultural landscapes in Nepal, Agr. Ecosyst. Environ., 333, 107973, https://doi.org/10.1016/j.agee.2022.107973, 2022.
Kéry, M. and Royle, J. A.: Hierarchical Bayes estimation of species richness and occupancy in spatially replicated surveys, J. Appl. Ecol., 45, 589–598, https://doi.org/10.1111/j.1365-2664.2007.01441.x, 2008.
Khamcha, D., Corlett, R. T., Powell, L. A., Savini, T., Lynam, A. J., and Gale, G. A.: Road induced edge effects on a forest bird community in tropical Asia, Avian Res., 9, 20, https://doi.org/10.1186/s40657-018-0112-y, 2018.
King, S., Elphick, C. S., Guadagnin, D., Taft, O., and Amano, T.: Effects of landscape features on waterbird use of rice fields, Waterbirds, 33, 151–159, https://doi.org/10.1675/063.033.s111, 2010.
Kouadja, S., Odoukpe, G., Konan, M., and Yaokokore-Beibro, H.: Composition and abundance of rice field birds in the District of Yamoussoukro (central Côte d'Ivoire), J. Appl. Biosci., 186, 19571–19587, 2023.
Lê, S., Josse, J., and Husson, F.: FactoMineR: An R package for multivariate analysis, J. Stat. Softw., 25, 1–18, https://doi.org/10.18637/jss.v025.i01, 2008.
Lee, M. B., Chen, D., Liu, F., and Zou, F.: Effects of spatial and temporal crop changes on bird diversity in peri-urban agricultural lands, Basic Appl. Ecol., 80, 138–145, https://doi.org/10.1016/j.baae.2024.09.007, 2024.
Leyequién, E., de Boer, W. F., and Toledo, V. M.: Bird community composition in a shaded coffee agro-ecological matrix in Puebla, Mexico: The effects of landscape heterogeneity at multiple spatial scales, Biotropica, 42, 236–245, https://doi.org/10.1111/j.1744-7429.2009.00553.x, 2010.
Longoni, V.: Rice fields and waterbirds in the Mediterranean region and the Middle East, Waterbirds, 33, 83–96, https://doi.org/10.1675/063.033.s106, 2010.
Lopes, P., Lacerda, I., Chaves, P., de Lima, R. F., Camará, M., Lecoq, M., Palmeirim, J. M., and Rainho, A.: Data and code for: Landscape context outweighs field-scale features in structuring bird communities in West African smallholder rice agroecosystems, OSF [data set/code], https://doi.org/10.17605/OSF.IO/TRFNQ, 2026.
Lu, X., Jia, Y., and Wang, Y.: The effect of landscape composition, complexity, and heterogeneity on bird richness: a systematic review and meta-analysis on a global scale, Landsc. Ecol., 39, https://doi.org/10.1007/s10980-024-01933-w, 2024.
Luther, D. A., Wolfe, J. D., Johnson, E., Stouffer, P. C., Batchelor, J., and Tarwater, C. E.: Habitat use of Amazonian birds varies by age and foraging guild along a disturbance gradient, P. R. Soc. B, 291, 20240866, https://doi.org/10.1098/rspb.2024.0866, 2024.
Marcolin, F., Lakatos, T., Gallé, R., and Batáry, P.: Fragment connectivity shapes bird communities through functional trait filtering in two types of grasslands, Global Ecol. Conserv., 28, e01687, https://doi.org/10.1016/j.gecco.2021.e01687, 2021.
Martin, E. A., Ratsimisetra, L., Laloë, F., and Carrière, S. M.: Conservation value for birds of traditionally managed isolated trees in an agricultural landscape of Madagascar, Biodivers. Conserv., 18, 2719–2742, https://doi.org/10.1007/s10531-009-9671-x, 2009.
Matuoka, M. A., Benchimol, M., Almeida-Rocha, J. M. de, and Morante-Filho, J. C.: Effects of anthropogenic disturbances on bird functional diversity: A global meta-analysis, Ecol. Indic., 116, 106471, https://doi.org/10.1016/j.ecolind.2020.106471, 2020.
McGarigal, K., Cushman, S., and Ene, E.: FRAGSTATS v4: Spatial Pattern Analysis Program for Categorical Maps, , https://www.fragstats.org (last access: 31 January 2025), 2023.
Miguet, P., Jackson, H. B., Jackson, N. D., Martin, A. E., and Fahrig, L.: What determines the spatial extent of landscape effects on species?, Landsc. Ecol., 31, 1177–1194, https://doi.org/10.1007/s10980-015-0314-1, 2016.
Nachuha, S.: Is waterbird distribution within rice paddies of eastern Uganda affected by the different stages of rice growing?, in: Proceedings of the 12th Pan-African Ornithological Congress, 2008, edited by: Harebottle, D. M., Craig. A. J. F. K., Anderson, M. D., Rakotomanana, H., and Muchai, M., Animal Demography Unit., Cape Town, 44–49, ISBN 978-0-79-922361-3, 2009.
Newbold, T., Boakes, E. H., Hill, S. L., Harfoot, M. B., and Collen, B.: The present and future effects of land use on ecological assemblages in tropical grasslands and savannas in Africa, Oikos, 126, 1760–1769, https://doi.org/10.1111/oik.04338, 2017.
Odoukpe, G. and Yaokokore-Beibro, H.: Avifaune des champs de riz de la zone humide de Grand-Bassam (Côte d'Ivoire), Int. J. Biol. Chem. Sci., 8, 1458–1480, https://doi.org/10.4314/ijbcs.v8i4.10, 2014.
Otieno, N. E. and Mukasi, J.: Tropical insectivorous birds' predation patterns that promote forest–farmland trophic connectivity for integrated top–down pest biocontrol, Front. Environ. Sci., 11, 1194267, https://doi.org/10.3389/fenvs.2023.1194267, 2023.
Perrings, C. and Halkos, G.: Agriculture and the threat to biodiversity in sub-saharan Africa, Environ. Res. Lett., 10, 095015, https://doi.org/10.1088/1748-9326/10/9/095015, 2015.
Priyadarshana, T. S., Martin, E. A., Sirami, C., Woodcock, B. A., Goodale, E., Martínez-Núñez, C., Lee, M. B., Pagani-Núñez, E., Raderschall, C. A., Brotons, L., Rege, A., Ouin, A., Tscharntke, T., and Slade, E. M.: Crop and landscape heterogeneity increase biodiversity in agricultural landscapes: A global review and meta-analysis, Ecol. Lett., 27, e14412, https://doi.org/10.1111/ele.14412, 2024.
Propper, C. R., Sedlock, J. L., Smedley, R. E., Frith, O., Shuman-Goodier, M. E., Grajal-Puche, A., Stuart, A. M., and Singleton, G. R.: Balancing food security, vertebrate biodiversity, and healthy rice agroecosystems in Southeast Asia, Crop Environ., 3, 43–50, https://doi.org/10.1016/j.crope.2023.11.005, 2024.
QGIS.org: QGIS Geographic Information System, https://qgis.org (last access: 20 April 2023), 2023.
R Core Team: R: A Language and Environment for Statistical Computing, https://www.r-project.org/ (last access: 20 April 2023), 2022a.
R Core Team: RStudio: Integrated Development Environment for R, http://www.rstudio.com/ (last access: 20 April 2023), 2022b.
Redlich, S., Martin, E. A., Wende, B., and Steffan-Dewenter, I.: Landscape heterogeneity rather than crop diversity mediates bird diversity in agricultural landscapes, PLOS ONE, 13, 1–14, https://doi.org/10.1371/journal.pone.0200438, 2018.
Renfrew, R., Ribic, C., and Nack, J.: Edge avoidance by nesting grassland birds: a futile strategy in a fragmented landscape, Auk, 122, 618–636, https://doi.org/10.1093/auk/122.2.618, 2005.
Rigal, S., Dakos, V., Alonso, H., Auniņš, A., Benkő, Z., Brotons, L., Chodkiewicz, T., Chylarecki, P., de Carli, E., del Moral, J. C., Doma̧, C., Escandell, V., Fontaine, B., Foppen, R., Gregory, R., Harris, S., Herrando, S., Husby, M., Ieronymidou, C., Jiguet, F., Kennedy, J., Klvaňová, A., Kmecl, P., Kuczyński, L., Kurlavičius, P., Kålås, J. A., Lehikoinen, A., Lindström, Å., Lorrillière, R., Moshøj, C., Nellis, R., Noble, D., Eskildsen, D. P., Paquet, J. Y., Pélissié, M., Pladevall, C., Portolou, D., Reif, J., Schmid, H., Seaman, B., Szabo, Z. D., Szép, T., Teufelbauer, N., Trautmann, S., van Turnhout, C., Vermouzek, Z., Vikstrøm, T., Voříšek, P., Weiserbs, A., Florenzano, G. T., and Devictor, V.: Farmland practices are driving bird population decline across Europe, P. Natl. Acad. Sci. USA, 120, e2216573120, https://doi.org/10.1073/pnas.2216573120, 2023.
Rolando, A.: On the ecology of home range in birds, Rev. Ecol. (Terre Vie), 57, 53–73, https://doi.org/10.3406/revec.2002.2381, 2002.
Şekercioglu, Ç. H., Mendenhall, C. D., Oviedo-Brenes, F., Horns, J. J., Ehrlich, P. R., and Daily, G. C.: Long-term declines in bird populations in tropical agricultural countryside, P. Natl. Acad. Sci. USA, 116, 9903–9912, https://doi.org/10.1073/pnas.1802732116, 2019.
Sólymos, P., Matsuoka, S. M., Stralberg, D., Barker, N. K. S., and Bayne, E. M.: Phylogeny and species traits predict bird detectability, Ecography, 41, 1595–1603, https://doi.org/10.1111/ecog.03415, 2018.
Son, S.-J., Choi, G., Do, M. S., and Nam, H.-K.: How do structural characteristics and landscape factors of rice paddies in Republic of Korea affect the species richness and abundance of birds utilizing them?, SSRN [preprint], https://doi.org/10.2139/ssrn.4987027, 2024.
Stafford, J. D., Kaminski, R. M., and Reinecke, K. J.: Avian foods, foraging and habitat conservation in world rice fields, Waterbirds, 33, 133–150, https://doi.org/10.1675/063.033.s110, 2010.
Stanton, R. L., Morrissey, C. A., and Clark, R. G.: Analysis of trends and agricultural drivers of farmland bird declines in North America: A review, Agr. Ecosyst. Environ., 254, 244–254, https://doi.org/10.1016/j.agee.2017.11.028, 2018.
Tabachnick, B. G., Fidell, L. S., and Ullman, J. B.: Using multivariate statistics, 7th edn., Pearson, Boston, ISBN 978-0-13-479054-1, 2007.
Tchetgen Tchetgen, E., Sun, B., and Walter, S.: The GENIUS approach to robust Mendelian randomization inference, Stat. Sci., 36, 443–464, https://doi.org/10.1214/20-STS802, 2021.
Temudo, M. P.: Planting Knowledge, Harvesting Agro-Biodiversity: A Case Study of Southern Guinea-Bissau Rice Farming, Hum. Ecol., 39, 309–321, https://doi.org/10.1007/s10745-011-9404-0, 2011.
The Climate Change Knowledge Portal: Guinea-Bissau, https://climateknowledgeportal.worldbank.org/country/guinea-bissau (last access: 27 March 2025).
Tobias, J. A., Sheard, C., Pigot, A. L., Devenish, A. J. M., Yang, J., Sayol, F., Neate-Clegg, M. H. C., Alioravainen, N., Weeks, T. L., Barber, R. A., Walkden, P. A., MacGregor, H. E. A., Jones, S. E. I., Vincent, C., Phillips, A. G., Marples, N. M., Montaño-Centellas, F. A., Leandro-Silva, V., Claramunt, S., Darski, B., Freeman, B. G., Bregman, T. P., Cooney, C. R., Hughes, E. C., Capp, E. J. R., Varley, Z. K., Friedman, N. R., Korntheuer, H., Corrales-Vargas, A., Trisos, C. H., Weeks, B. C., Hanz, D. M., Töpfer, T., Bravo, G. A., Remeš, V., Nowak, L., Carneiro, L. S., Moncada R., A. J., Matysioková, B., Baldassarre, D. T., Martínez-Salinas, A., Wolfe, J. D., Chapman, P. M., Daly, B. G., Sorensen, M. C., Neu, A., Ford, M. A., Mayhew, R. J., Fabio Silveira, L., Kelly, D. J., Annorbah, N. N. D., Pollock, H. S., Grabowska-Zhang, A. M., McEntee, J. P., Carlos T. Gonzalez, J., Meneses, C. G., Muñoz, M. C., Powell, L. L., Jamie, G. A., Matthews, T. J., Johnson, O., Brito, G. R. R., Zyskowski, K., Crates, R., Harvey, M. G., Jurado Zevallos, M., Hosner, P. A., Bradfer-Lawrence, T., Maley, J. M., Stiles, F. G., Lima, H. S., Provost, K. L., Chibesa, M., Mashao, M., Howard, J. T., Mlamba, E., Chua, M. A. H., Li, B., Gómez, M. I., García, N. C., Päckert, M., Fuchs, J., Ali, J. R., Derryberry, E. P., Carlson, M. L., Urriza, R. C., Brzeski, K. E., Prawiradilaga, D. M., Rayner, M. J., Miller, E. T., Bowie, R. C. K., Lafontaine, R. M., Scofield, R. P., Lou, Y., Somarathna, L., Lepage, D., Illif, M., Neuschulz, E. L., Templin, M., Dehling, D. M., Cooper, J. C., Pauwels, O. S. G., Analuddin, K., Fjeldså, J., Seddon, N., Sweet, P. R., DeClerck, F. A. J., Naka, L. N., Brawn, J. D., Aleixo, A., Böhning-Gaese, K., Rahbek, C., Fritz, S. A., Thomas, G. H., and Schleuning, M.: AVONET: morphological, ecological and geographical data for all birds, Ecol. Lett., 25, 581–597, https://doi.org/10.1111/ele.13898, 2022.
Tong, Y. D.: Rice Intensive Cropping and Balanced Cropping in the Mekong Delta, Vietnam – Economic and Ecological Considerations, Ecol. Econ., 132, 205–212, https://doi.org/10.1016/j.ecolecon.2016.10.013, 2017.
Tubiello, F. N., Conchedda, G., Casse, L., Hao, P., De Santis, G., and Chen, Z.: A new cropland area database by country circa 2020, Earth Syst. Sci. Data, 15, 4997–5015, https://doi.org/10.5194/essd-15-4997-2023, 2023.
Tscharntke, T., Sekercioglu, C. H., Dietsch, T. V., Sodhi, N. S., Hoehn, P., and Tylianakis, J. M.: Landscape constraints on functional diversity of birds and insects in tropical agroecosystems, Ecology, 89, 944–951, https://doi.org/10.1890/07-0455.1, 2008.
Tscharntke, T., Tylianakis, J. M., Rand, T. A., Didham, R. K., Fahrig, L., Batáry, P., Bengtsson, J., Clough, Y., Crist, T. O., Dormann, C. F., Ewers, R. M., Fründ, J., Holt, R. D., Holzschuh, A., Klein, A. M., Kleijn, D., Kremen, C., Landis, D. A., Laurance, W., Lindenmayer, D., Scherber, C., Sodhi, N., Steffan-Dewenter, I., Thies, C., van der Putten, W. H., and Westphal, C.: Landscape moderation of biodiversity patterns and processes – eight hypotheses, Biol. Rev., 87, 661–685, https://doi.org/10.1111/j.1469-185X.2011.00216.x, 2012.
van Osta, J. M., Dreis, B., Grogan, L. F., and Castley, J. G.: Local resource availability drives habitat use by a threatened avian granivore in savanna woodlands, PLOS ONE, 19, e0306842, https://doi.org/10.1371/journal.pone.0306842, 2024.
Voudouri, A., Chaideftou, E., and Sfougaris, A.: Topsoil seed bank as feeding ground for farmland birds: A comparative assessment in agricultural habitats, Land, 10, 967, https://doi.org/10.3390/land10090967, 2021.
Vukeya, L. R., Mokotjomela, T. M., and Pillay, N.: Determining the status of ecosystem degradation trends and their implications for ecological integrity in the southern African grassland biome, Global Ecol. Conserv., 58, e03462, https://doi.org/10.1016/j.gecco.2025.e03462, 2025.
Weller, M.: Wetland birds: habitat resources and conservation implications, Cambridge University Press, Cambridge, UK, ISBN 978-0-52-163362-8, 1999.
Wright, H. L., Lake, I. R., and Dolman, P. M.: Agriculture-a key element for conservation in the developing world, Conserv. Lett., 5, 11–19, https://doi.org/10.1111/j.1755-263X.2011.00208.x, 2012.
Wymenga, E. and Zwarts, L.: Use of rice fields by birds in West Africa, Waterbirds, 33, 97–104, https://doi.org/10.1675/063.033.s107, 2010.
Yamaura, Y., Connor, E. F., Royle, J. A., Itoh, K., Sato, K., Taki, H., and Mishima, Y.: Estimating species–area relationships by modeling abundance and frequency subject to incomplete sampling, Ecol. Evol., 6, 4836–4848, https://doi.org/10.1002/ece3.2244, 2016.
Yaokororé-Béibro, H. and Odoukpé, G.: Dynamic of waterbirds on rice-fields in the Grand-Bassam wetlands (Ivory Coast, West Africa), Alauda, 83, 255–262, 2015.
Zhu, Y., Liu, Y., Sheng, S., Zheng, J., Wu, S., Cao, Z., Zhang, K., and Xu, Y.: Quantifying the effects of landscape and habitat characteristics on structuring bird assemblages in urban habitat patches, Sci. Rep., 14, 12707, https://doi.org/10.1038/s41598-024-63333-z, 2024.
Zwarts, L., Bijlsma, R. G., van der Kamp, J., and Wymenga, E.: Living on the Edge Wetlands and Birds in a Changing Sahel, 2nd edn., KNNV Publishing, Zeist, the Netherlands, ISBN 978-9-05-011280-2, 2009.
Zwarts, L., Bijlsma, R. G., and van der Kamp, J.: Selection by birds of shrub and tree species in the Sahel, Ardea, 111, 143–174, https://doi.org/10.5253/arde.2022.a20, 2023.